Филогенез в онтогенезе что это
Лекция «Возрастная анатомия опорно-двигательного аппарата»
Стадии развития скелета в филогенезе.

У животных выделяют наружный и внутренний скелет.
Наружный скелет у разных животных (рис. 1) имеет разное строение и происхождение. У многих беспозвоночных он является продуктом выделения кожного эпителия: кутикула дождевого червя, хитин членистоногих, известковые раковины молюсков.
Наружный скелет у позвоночных появляется в форме чешуи у рыб. Из чешуй у высших рыб развиваются покровные кости головы и плечевого пояса.
Чешуя рыб и кожные окостенения наземных позвоночных всегда дополняются внутренним скелетом.
Внутренний скелет у низших животных (рис. 1) развит слабо и представляет собой систему соединительнотканных образований, иногда включающих рогоподобные волокна, кремниевые или известковые иглы.
Внутренний скелет у головоногих молюсков представлен хрящом.
У позвоночных животных внутренний скелет всегда хорошо развит.
У бесчерепных он перепончатый, у низших рыб – хрящевой, у высших рыб и наземных позвоночных он построен преимущественно из костной ткани.
Развитие скелета в онтогенезе у человека.

Согласно основному биогенетическому закону Геккеля-Мюллера онтогенез есть краткое повторение филогенеза. Онтогенез твердого скелета у человека не является исключением: в развитии костей у человека выделяются три последовательных стадии (рис. 2):
1. Соединительнотканная.
2. Хрящевая.
3. Костная.
Большинство костей в своем развитии последовательно проходят все три стадии – это вторичные кости. Ряд костей при развитии пропускают хрящевую стадию – это первичные кости. К первичным по развитию костям относятся: кости свода черепа, кости лицевого черепа, часть ключицы (акромиальный конец).
Первичные и вторичные кости.

По развитию кости человека делятся на две группы (рис. 3):
Характеристика остеобластов и остеокластов развиваюшейся кости.

Для развития костной ткани в костях необходимо наличие популяций двух видов клеток (рис. 4):
Остеобласты представляют собой кубовидной формы клетки (20-30 мкм в диаметре) с одним крупным ядром, располагающиеся близко друг к другу на костном матриксе (межклеточном веществе). Фибробласты продуцируют все компоненты костного матрикса. Они имеют два разных эмбриональных источника:
Для формирования кости как органа необходимо совместная работа двух видов клеток: остеобластов и остеокластов.
Cпособы развития костей (окостенения).


В зависимости от того где начинается формирование костной ткани в костях (включая их закладки) выделяют четыре способа окостенения (рис. 5):
При эндесмальном окостенении (рис. 5) первичная точка окостенения появляется в центре соединительнотканной закладки кости. Затем новообразующаяся костная ткань распространяется от цента органа к периферии. Таким способом окостеневают первичные кости. На месте первичной точки окостенения обычно наблюдается утолщение (например, теменной бугор, наружный затылочный выступ и т.п.).
Периходральное окостенение характерно для вторичных костей. Остеобласты выстраиваются на поверхности хрящевой закладки кости и начинают синтезировать костный матрикс. Это приводит с сдавливанию и нарушению трофика подлежащей хрящевой ткани, изменения которой активирует остеокласты. В результате этого на поверхности хрящевой закладки кости появляется и постепенно нарастает костная ткань (рис. 5). За счет перихондрального окостенения формируется компактное костное вещество. У длинных трубчатых костей так во внутриутробном периоде образуется диафиз.
При энхондральном окостенении точка (первичный очаг) окостенения появляется в центре хрящевой закладки кости. Затем костная ткань разрастается из центра к периферии (рис. 6). В результате этого формируется губчатое костное вещество. Этим способом развиваются вторичные кости: эпифизы и апофизы трубчатых костей, губчатые, плоские (кроме свода черепа) кости.
Периостальное окостенение происходит за счет надкостницы (periosteum, лат – надкостница). У детей за счет надкостницы кости растут в толщину (напоминаем, что рост кости в длину идет за счет метафизарного хряща)(рис. 6). У взрослых периостальное окостенение обеспечивает физиологическую регенерацию кости.
Развитие костей туловища (общие свойства). Развитие и аномалии развития позвонков.


Рис. 8. Развитие и аномалии развития позвонков.

Рис. 9. Расщелина дуг позвонков на протяжении всех грудных позвонков.
Кости туловища по развитию относятся к вторичным костям. Они окостеневают энхондрально (рис. 7).
Развитие позвонков:
У зародыша закладывается 38 позвонков: 7 шейных, 13 грудных, 5 поясничных, 12-13 крестцовых и копчиковых (рис. 8).
13-й грудной превращается в 1-й поясничный, последний поясничный – в 1-й крестцовый, Идет редукция большинства копчиковых позвонков.
Каждый позвонок имеет первоначально три ядра окостенения: в теле и по одному в каждой половинке дуги. Они срастаются лишь к третьему году жизни.
Вторичные центры появляются по верхнему и нижнему краям тела позвонка у девочек в 6-8 лет, у мальчиков – в 7-9 лет. Они прирастают к телу позвонка в 20-25 лет.
Самостоятельные ядра окостенения образуются в отростках позвонков.
Аномалии развития позвонков (рис. 8, 9):
— Врожденные расщелины позвонков:
— Клиновидные позвонки и полупозвонки.
— Платиспондилия – расширение тела позвонка в поперечнике.
— Брахиспондилия – уменьшение тела позвонка по высоте, уплощение и укорочение.
— Аномалии суставных отростков: аномалии положения, аномалии величины, аномалии сочленения, отсутствие суставных отростков.
— Спондилолиз – дефект в межсуставной части дуги позвонка.
— Врожденные синостозы: полный и частичный.
— Os odontoideum – неслияние зуба с телом осевого позвонка.
— Ассимиляция (окципитализация) атланта – слияние атланта с затылочной костью.
— Сакрализация – полное или частичное слияние последнего поясничного позвонка с крестцом.
— Люмбализация – наличие шестого поясничного позвонка (за счет мобилизации первого крестцового).
Развитие и аномалии развития ребер и грудины.

Рис. 10. Развитие и аномалии развития ребер.

Рис. 11. Развитие и аномалии развития грудины.
Развитие ребер (рис. 10):
Закладывается 13 пар ребер. Затем 13-е ребро редуцируется и срастается с поперечным отростком 1-го поясничного позвонка.
Основных точек окостенения в ребре две: точка окостенения на месте будущего угла ребра (окостеневает тело ребра) и в головке ребра (на 15-20 году жизни). У 10 верхних ребер появляется точка окостенения в бугорке ребра.
Передние концы 9 пар верхних ребер образуют грудные полоски – источник развития грудины.
Развитие грудины (рис. 11):
Источником развития грудины являются грудные полоски – расширенные концы хрящевых концов девяти пар верхних ребер. В грудине бывает до 13 точек окостенения.
Аномалии развития ребер (рис. 10):
— Отсутствие ребра
— Отсутствие части ребра
— Дефект ребра
— Раздвоение ребра (вилка Лушки)
— Шейное ребро
— XIII ребро
Аномалии развития грудины (рис. 11):
— Аплазия рукоятки грудины
— Отсутствие отдельных сегментов тела грудины <
— Расщепление грудину
— Отсутствие тела грудины
— Воронкообразная деформация
— Куриная грудь
Развитие костей конечностей.



Рис. 14. Развитие эпифизов трубчатых костей.

Рис. 15. Развитие костей верхней конечности.

Рис. 16. Развитие тазовой и бедренной костей.


Кости конечностей по развитию относятся к вторичным костям. Исключение представляет собой ключица: ее тело и акромиальный конец окостеневают эндесмально (точка окостенения появляется на 6-7-й неделях внутриутробного развития.
Диафизы длинных трубчатых костей окостеневают перихондральными и энходральными способами. В диафизах первичная точка окостенения появляется на 2-м – начале 3-го месяцев внутриутробного развития и растет по направлению к проксимального и дистальному эпифизам.
Эпифизы и апофизы длинных трубчатых костей окостеневают энходральным способом. Они у новорожденных хрящевые. Вторичные точки окостенения появляются в течение первых 5-10 лет жизни. Исключение составляют эпифизы костей, образующих коленный сустав: точка окостенения в дистальном конце бедренной кости появляется на 6 месяце, а в проксимальном конце большеберцовой кости – на 7 месяце внутриутробного развития. Прирастают эпифизы к диафизам после 15-17 лет и позже.
Варианты и аномалии развитие костей конечностей.

Рис. 19. Аномалии развития костей верхней конечности.

Рис. 20. Аномалии развития костей нижней конечности.
Аномалии развития лопатки:
Аномалии развития ключицы:
Варианты и аномалии развития плечевой кости
Аномалии развития костей предплечья:
Аномалии развития костей кисти:
Варианты и аномалии развития тазовой кости:
Варианты и аномалии развития бедренной кости:
Варианты и аномалии развития костей голени:
Варианты и аномалии развития костей стопы
Развитие костей черепа.


Рис. 22. Источники развития костей лицевого черепа.


Рис. 24. Развитие костей черепа после рождения.
Кости свода и лицевого черепа по развитию относятся к первичным костям, окостеневающим на основе соединительной ткани эндесмальным способом окостенения.
Кости лицевого черепа развиваются на основе жаберных дуг (первой и второй висцеральной дуги).
Из первой висцеральной дуги развиваются следующие кости: верхняя, нижняя челюсти, частично скуловая и небные кости, медиальная пластинка крыловидного отростка клиновидной кости; молоточек, наковальня – слуховые косточки; костное небо и его швы, нижняя часть глазницы.
Из второй висцеральной дуги развиваются: стремечко, шиловидный отросток височной кости, малые рога подъязычной.
Кости основания черепа проходят три стадии развития: соединительнотканную, хрящевую и костную. Т.е. они являются вторичными. Они окостеневают энхондрально.
Варианты и аномалии развития костей черепа.

Рис. 25. Вставочные кости швов черепа (слева), деформации черепа (справа).


Известны следующие аномалии развития черепа
Филогенез соединений костей



У рыб, обитающих в водной среде, многочисленные кости скелета (рис. 28) соединяются при помощи непрерывных соединений: соеденительнотканных и хрящевых.
Важным биомеханическим фактором, повлиявшим на филогенез соединений костей, является выход животных на сушу. Кратковременное пребываниена твердой поверхности (в том числе перемещение с места на место), которое наблюдается, напрмер, у двоякодышащих рыб, приводит к появлению гемиартрозов между костями конечностей (рис. 29). Большинство исследователей считают такую форму пререходной от снартроза к диартрозу.
Окончательный выход животных на сушу формирует два направления морфогенеза мягкого остова. Во-первых, формируются суставы со всеми обязательными и вспомогательными элементами и высокой степенью подвижности. Во-вторых, в местах контакта костей с увеличившейся нагрузкой (из-за возросшего действия силы тяжести) формируются синостозы (кости срастаются между собой). Признаки обоих изменений строения соединений костей уже выявляются у земноводных (рис. 30).
Онтогенез соединений костей

Рис. 31. Варианты дисплазии тазобедренного сустава.
В онтегенезе соединения костей наблюдаются сходные с филогенезом тенденции. Первоначально все соединения образованые скоплением мезенхимальных клеток (эмбриональная соединительная ткань).
В конце первой половины пренатального онтогенеза (16-18-ая недели внутриутробного развития) между костями, которые смещаются (движутся) друг относительно друга, формируются суставы. Важным биомеханическим фактором их морфогенеза являются силы мышц, двигающих кости. Между зачатками костей, которые соединяются без смещание, формируются непрерывные соединения.
У новорожденных имеется закладка всех элементов суставов на нижней конечности. Однако большинство из них достигают функциональной зрелости к юношескому возрасту.
Основной аномалией развития соединения костей является дисплазия суставов. Для данные аномалии характерно изменение формы одной из суставных поверхностей, сопровождающееся изменениями строения расположенных рядом элементов сустава (рис. 31).
ПОНЯТИЕ ОНТОГЕНЕЗА И ФИЛОГЕНЕЗА, ИХ ВЗАИМОСВЯЗИ
Для выяснения роли адаптации в филогенезе и онтогенезе необходимо вспомнить эти понятия.
Термин филогенез ввел Э. Геккель в 1866 году. Филогенез изучает историю возникновения и развития вида (Phyle – племя, genesis – развитие). В более широком плане филогенез отражает историческое развитие живых организмов на Земле. Раздел биологии, изучающий филогенез, называется филогенетикой. Термин «филогенетический» часто используют как синоним термина «эволюционный». Но следует иметь в виду, что филогенетика не изучает механизмы эволюции, а лишь констатирует родственные связи между таксонами (от царств до видов).
Выражение «филогенетические преобразования» следует понимать как преобразования в ходе исторического развития группы организмов. В современной филогенетике используют данные не только палеонтологии ( по Геккелю), но и генетики, биохимии, молекулярной биологии, этологии, биогеографии и физиологии. Последнее в большей степени интересует нас.
Графическое изображение филогенеза – родословное (филогенетическое) древо. Основной движущей силой, определяющей адаптивный характер филогенетических преобразований организмов, является естественный отбор. Конкретные направления филогенеза ограничиваются исторически сложившимися особенностями генетической системы, морфогенеза и фенотипа каждой конкретной группы.
Индивидуальное развитие организма называется онтогенез. К. Э. Бэр в 1828 г. сформулировал закон индивидуального развития, в котором наряду с планами строения организма выделил отвечающие им схемы развития. Его интересовал вопрос, как строится организм? Онтогенез он рассматривал как процесс построения организма. В основные положения закона индивидуального развития К. Э. Бэра вошли:
1. Общее каждой более крупной животной группы образуется в зародыше раньше, чем специальное.
2. Из более общего в области отношения форм образуется менее общее и так далее, пока, наконец, не возникает самое специальное.
3. Зародыш каждой животной формы отнюдь не повторяет при развитии другие животные формы, а, напротив, скорее обосабливается от них.
4. В основе своей, значит, что зародыш высшей животной формы никогда не бывает подобен другой форме, а лишь ее зародышу.
В итоге К. Э. Бэр делает заключение о том, что история развития особи есть история растущей во всех отношениях индивидуальности. В ходе онтогенеза происходит преобразование её морфофизиологических, физиолого-биохимических и цитогенетических признаков.
Таким образом, онтогенез– это индивидуальное развитие opганизма, в ходе которого происходит преобразование его морфофизиологических, физиолого-биохимических и цитогенетических признаков. Онтогенез включает две группы процессов: морфогенез и воспроизведение (репродукцию). В результате морфогенеза формируется репродуктивно зрелая особь. Онтогенез характеризуется устойчивостью – гомеорезом. Гомеорез – это стабилизированный поток событий, который представляет собой процесс реализации генетической программы строения, развития и функционирования организма.
Основные атрибуты онтогенеза:
– исходная запрограммированность процессов. Наличие уникальной неизменной генетической программы развития, сформированной вследствие мейоза и оплодотворения;
– необратимость онтогенеза. При реализации генетической программы невозможен возврат к предыдущим стадиям;
– углубление специализации: по мере развития уменьшается вероятность смены траектории онтогенеза;
– адаптивный характер: поливариантность онтогенеза обеспечивает возможность приспособления к различным условиям;
– неравномерность темпов: скорость процессов роста и развития изменяется;
– целостность и преемственность отдельных этапов. Признаки, появляющиеся на более поздних стадиях, базируются на признаках, проявляющихся на ранних стадиях;
– наличие цикличности: существует цикличность старения и омоложения;
– наличие критических периодов, связанных с выбором пути в узловых точках (точках бифуркации) или с преодолением энергетических порогов.
Основные типы онтогенеза:
1. Онтогенез организмов с бесполым размножением и/или при зиготном мейозе (прокариоты и некоторые эукариоты).
2. Онтогенез организмов с чередованием ядерных фаз при споровом мейозе (большинство растений и грибов).
4. Онтогенез с наличием личиночных и промежуточных стадий: от первично-личиночного анаморфоза до полного метаморфоза. При недостатке питательных веществ в яйце личиночные стадии позволяют завершить морфогенез, а также в ряде случаев обеспечивают расселение особей.
5. Онтогенез с выпадением отдельных стадий. Утрата личиночных стадий и/или стадий бесполого размножения. Утрата конечных стадий и размножение на ранних этапах онтогенеза: неотения.
Дата добавления: 2017-02-20 ; просмотров: 14986 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Онтогенез и филогенез
Урок 2. Подготовка к ЕГЭ по биологии


В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобрев в каталоге.
Получите невероятные возможности



Конспект урока «Онтогенез и филогенез»
Процесс индивидуального развития особи от начала её существования и до конца жизни называют онтогенезом.
У бактерий и простейших онтогенез практически совпадает с клеточным циклом.
У многоклеточных видов, которые размножаются бесполым путём, онтогенез начинается с момента выделения группы клеток материнского организма, которые затем делятся митозом и формируют новую особь со всеми системами и органами.
У тех видов, которые размножаются половым путём, онтогенез начинается с момента оплодотворения яйцеклетки и образования зиготы — первой клетки нового организма.
У животных выделяют три типа онтогенеза: личиночный, яйцекладный и внутриутробного развития.
Личиночный тип развития.
Он встречается, например, у насекомых, рыб, земноводных. Желтка в яйцеклетках этих организмов мало, и зигота быстро развивается в личинку, которая самостоятельно питается и растёт. Затем, по прошествии какого-то времени, происходит метаморфоз, — то есть превращение личинки во взрослую особь.
У некоторых видов наблюдается даже целая цепочка превращений из одной личинки в другую и только потом — во взрослую особь.
Например, у кузнечика личинка питается, растёт, несколько раз линяет. После окончательной линьки она становится взрослой половозрелой особью с крыльями и больше не растёт.
Яйцекладный тип онтогенеза
Он наблюдается у рептилий, птиц и яйцекладущих млекопитающих, яйцеклетки которых богаты желтком. Зародыш таких видов развивается внутри яйца; личиночная стадия отсутствует.
Питание зародыша обеспечивает желток. Содержимое яйца окружено двумя подскорлупковыми оболочками, внутренней и наружной. Снаружи находится скорлупа. После откладки яйца на его тупом конце постепенно образуется воздушная камера.
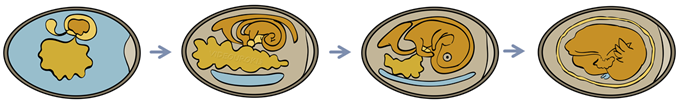
Однако у большинства млекопитающих яйцо не образуется, а зародыш развивается внутри тела самки в половом органе — матке.
Внутриутробный тип онтогенеза
Он наблюдается у большинства млекопитающих, в том числе и у человека. При этом развивающийся зародыш задерживается в материнском организме, образуется временный орган — плацента, через который организм матери обеспечивает все потребности растущего эмбриона: дыхание, питание, выделение и др. Внутриутробное развитие оканчивается процессом деторождения.
Любой вид индивидуального развития у многоклеточных животных принято делить на два периода: эмбриональный и постэмбриональный.
Эмбриональный период начинается с оплодотворения и представляет собой процесс формирования сложного многоклеточного организма, в котором представлены все системы органов.
Заканчивается этот период выходом личинки из своих оболочек (при личиночном типе онтогенеза), выходом особи из яйца (при яйцекладном типе) или рождением особи (при внутриутробном типе).
Постэмбриональный период начинается с завершения эмбрионального. Он включает в себя половое созревание, взрослое состояние, старение, которое заканчивается смертью.
Этапы эмбрионального развития на примере ланцетника.
Вспомним, что ланцетник ― это примитивное морское животное из семейства ланцетниковых. Ланцетник рассматривается как промежуточное звено между позвоночными и беспозвоночными животными.
В результате оплодотворения гаплоидные ядра половых клеток сперматозоида и яйцеклетки сливаются. Так возникает одноклеточная стадия развития организма — зигота, которая содержит диплоидный набор хромосом.
Зигота является началом нового организма. Она делится митозом.
Митотические деления называются делениями дробления.
Дробление (бластуляция) ― это процесс образования многоклеточного зародыша.
Первое деление происходит в вертикальной плоскости, и клетка делится на две одинаковые клетки с диплоидным набором хромосом. Их называют бластомерами.
Бластомеры не расходятся, а делятся ещё раз, и образуется уже 4 клетки.
Третье деление происходит в горизонтальной плоскости, и из четырёх образуется 8 бластомеров.
Далее продольные и поперечные деления сменяют друг друга. Возникает все больше бластомеров, которые не успевают расти.
После нескольких делений, когда число бластомеров достигает 32, они образуют полый шарик со стенками в один ряд клеток. Этот шарик получил название бластулы.
Бластула имеет внутри полость (пустое пространство). Эту полость называют первичной полостью тела или бластоцелью.
У человека на шестой день после оплодотворения бластула выходит из яйцевода в полость матки, а затем внедряется в её стенку. Этот процесс называется имплантацией зародыша.
После этого на одном из полюсов бластулы её клетки начинают делится быстрее, чем на другом, и впячиваться внутрь бластоцели. Этот процесс получил название гаструляции.
Так формируется второй внутренний слой клеток ― зародышевый листок. Такой двуслойный шарик называется гаструлой.
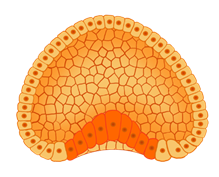
Гаструла состоит из двух зародышевых листков. Верхние клетки составляют наружный зародышевый листок ― эктодерму.
А внутренние ― внутренний зародышевый листок ― энтодерму.
Полость, которая образовалась внутри гаструлы, является первичной кишкой.
А отверстие, ведущее в первичную кишку, — первичным ртом.
У позвоночных животных на месте первичного рта в процессе эмбриогенеза образуется анальное отверстие, а настоящий (вторичный рот) возникает на противоположном полюсе зародыша. Поэтому млекопитающих (как и всех хордовых животных) относят к вторичноротым.
Между энтодермой и эктодермой образуется третий (средний) зародышевый листок — мезодерма.
Следующая стадия развития зародыша называется нейрулой.
На этой стадии происходит формирование нервной трубки и хорды.
У большинства хордовых хорда присутствует только в период эмбрионального развития, в дальнейшем замещаясь позвоночником.
У низших хордовых она сохраняется всю жизнь, выполняя опорную функцию, например, как у ланцетника.
На будущей спинной стороне зародыша начинается впячивание эктодермы в форме желобка — закладывается нервная пластинка, которая в дальнейшем развивается в нервную трубку.
Процесс образования нервной пластинки и её замыкание в нервную трубку в процессе зародышевого развития хордовых называется — нейруляцией.
Нервная трубка погружается под эктодерму и образует зачаток спинного мозга. А из его переднего широкого конца развивается головной мозг.
Таким образом, уже на ранних стадиях эмбрионального периода онтогенеза из внешне одинаковых бластомеров развиваются различные по строению и функциям ткани, органы и системы. Этот процесс получил название дифференцировки клеток.
Дифференцировка происходит благодаря активации определённых групп генов в различных клетках зародыша, благодаря чему в клетках синтезируются белки, каждый из которых будет выполнять те или иные функции.
Следом за эмбриональным развитием сразу наступает постэмбриональное развитие.
У разных организмов этот период может происходить по-разному.
Постэмбриональное развитие начинается с выхода новой особи из яйцевых оболочек (при живорождении) из организма матери.
Постэмбриональное развитие подразделяется на три периода:
ювенИльный период, продолжается до окончания полового созревания;
пубертАтный (период половой зрелости) и период старения.
Развитее организма в ювенильный период.
У разных видов развитие организма в ювенильный период может протекать по двум разным путям.
У одних развитие может быть прямое. У других непрямое.
Прямое развитие происходит без превращений. В этом случае вновь появившийся на свет организм отличается от взрослой особи только размерами, пропорциями и недоразвитием некоторых органов.
Такое развитие наблюдается у рыб, пресмыкающихся, птиц и млекопитающих.
У большинства млекопитающих зародыш развивается внутри тела самки. Такой вариант развития называют внутриутробным.
Когда организм новой особи становится способным к самостоятельному дыханию и питанию, происходят роды — из половых путей самки выходит детёныш.
Непрямое постэмбриональное развитие иногда требует сложной перестройки при переходе к взрослой форме.
Различают два типа непрямого развития — с полным и неполным превращением.
Развитие с превращением характерно для ряда насекомых и земноводных.
Личинки насекомых с неполным превращением внешне сходны со взрослыми особями.
Таким образом, при неполном превращении насекомое проходит три стадии: яйцо, личинка и имаго ― взрослое насекомое.
У насекомых при развитии с полным превращением особь проходит несколько последовательных стадий, отличающихся друг от друга образом жизни и характером питания.
При развитии с превращением из яйца появляется личинка, совершенно не похожая на взрослый организм. Такое развитие называется непрямым или развитием с метаморфозом, т. е. постепенным превращением организма во взрослую особь.
Таким образом, у насекомых с полным развитием в онтогенезе присутствуют четыре стадии: яйцо, личинка, куколка и имаго.
Ювенильный период всегда сопровождается ростом организма. Процесс роста запрограммирован генетически, а также здесь немалую роль играют условия существования.
Пубертатный период — период половой зрелости, у большинства позвоночных животных занимает, большую часть жизни.
После пубертатного периода начинаются изменения, которые снижают возможность организма к приспособлениям к изменяющимся условиям окружающей среды.

Филогенез — это историческое развитие организмов. Филогенез рассматривает эволюцию в качестве процесса, в котором генетическая линия — организмы от предка к потомкам — разветвляется во времени, и её отдельные ветви могут приобретать те или иные изменения или исчезать в результате вымирания. Этот процесс принято изображать графически в виде филогенетического древа, показывающего возможные родственные связи между отдельными ветвями живого.



