Какое количество структурных элементов у коры больших полушарий
Кора представлена слоем серого вещества толщиной 3-5 мм. В коре насчитывают до 15 и более млрд. нейронов, а число глиоцитов в головном мозге — более 100 млрд.
Развитие. Формирование коры больших полушарий происходит путем закономерной миграции нейробластов эпендимного слоя вдоль вертикально ориентированных радиальных глиоцитов. Первыми возникают наиболее поверхностный и глубокий слои коры. Затем возникают очередные последовательные волны миграции групп нейробластов, которые дифференцируются в нейроны V-гo, затем IV-гo слоя и д. т. Таким образом, нейробласты очередной волны миграции преодолевают слой нейронов, возникших от более ранней волны миграции. Это создает послойную (экранную) цитоархитектонику коры большого мозга.
Между нейронами устанавливаются сложные взаимосвязи в соответствии с их местом в составе рефлекторных дуг. Формируются ядерные и экранные нервные центры. Тесные взаимоотношения в ходе гистогенеза складываются между нейронами и клетками глии.
Строение. Все нейроны коры мулътиполярные. Среди них по форме клеток различают пирамидные и непирамидные (звездчатые, корзинчатые, веретенообразные, паукообразные и горизонтальные) нейроны. Пирамидные нейроны, наиболее характерны для коры, имеют тело, по форме напоминающее пирамиду, вершина которой обращена к поверхности коры.
От основания пирамидной клетки отходит аксон, имеющий коллатерали (возвратные, горизонтальные, косые). От вершины и боковых поверхностей тела отходят длинные дендриты (апикальные и базальные). Апикальные дендриты группы нейронов объединяются в дендритические пучки. На поверхности дендритов одного пирамидного нейрона может быть до 4-6 тыс. специальных рецепторных аппаратов — шипиков. Наличие в последних актомиозинового комплекса позволяет изменять площадь синаптического контакта, следовательно, влиять на синаптическую связь.
Величина тела пирамидных клеток варьирует от 10 до 150 мкм. Различают малые, средние, крупные и гигантские пирамиды. Пирамидные клетки — это эфферентные нейроны коры, их аксонные коллатерали образуют 3/4 всех синапсов в коре.
Звездчатые нейроны имеют тело в форме звезды. Дендриты отходят во все стороны от тела звездчатого нейрона. Они в большинстве случаев короткие и лишены шипиков. Аксоны звездчатых клеток образуют сложные разветвления около клетки. Это так называемые околоклеточные паутинообразные аксонные сети. Эти клетки встречаются в нижних слоях коры.
Корзинчатые клетки (мелкие и крупные), расположены во П-м и Ш-м слоях коры, своими многочисленными отростками образуют синаптические связи с телами пирамидных нейронов V-ro слоя. Клетки содержат медиатор (ГАМК), который тормозит передачу возбуждения.
Нейроглиоморфные клетки встречаются во всех слоях коры. Это мелкие мультиполярные нейроны с короткими ветвящимися денд-ритами и аксонами.
Биполярные нейроны — малочисленная группа клеток, от тела которых отходят аксон и дендрит. В целом соотношение между пирамидными и другими формами нейронов равно 85:15, то есть в пользу пирамидных нейронов.
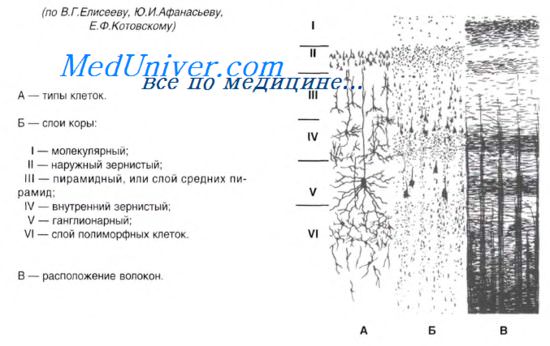
Цитоархитектоника. В двигательной зоне коры различают шесть основных слоев: молекулярный, наружный зернистый, пирамидный, внутренний зернистый, ганглионарный, слой полиморфных клеток.
В первом (наружном) молекулярном слое почти нет тел нейронов. Обнаруживаются единичные горизонтально ориентированные нейроны, тангенциальные ветвления нервных волокон нижележащих нейронов и глиальные клетки.
Второй, или наружный зернистый, слой содержит мелкие звездчатые и пирамидные нейроны размером около 10 мкм. Аксоны этих нейронов оканчиваются в III-м, IV-м и VI-м слоях коры, а дендриты поднимаются в молекулярный слой.
Третий слой — это слой средних и крупных пирамидных нейронов. Аксоны этих клеток образуют ассоциативные нервные волокна, идущие через белое вещество и соединяющие соседние участки коры.
Четвертый, или внутренний зернистый, слой содержит в основном мелкие звездчатые нейроны. Аксоны этих клеток, разветвляются в пределах соседних как выше, так и ниже лежащих слоев коры. Этот слой сильно развит в зрительной и слуховой зонах коры. Он состоит из сенсорных звездчатых нейронов, имеющих многочисленные ассоциативные связи с нейронами других типов.
Пятый — ганглионарный — слой образован большими пирамидными нейронами (клетками Беца). Апикальные дендриты нейронов направляются в молекулярный слой. Аксоны этих клеток идут в белое вещество, образуя комиссуральные и проекционные нервные волокна, и прежде всего пирамидные пути.
Шестой слой — слой полиморфных нейронов — содержит также много эфферентных пирамидных нейронов. Кроме того, здесь есть веретенообразные нейроны. Дендриты нейронов шестого слоя пронизывают всю толщину коры, достигая молекулярного слоя.
3.6.2. Структурно-функциональная организация коры больших полушарий
Кора больших полушарий (неокортекс) представлена скоплением нервных клеток, имеет многослойное строение (состоит из 6 слоев), множество борозд и извилин, за счет которых создается большая ее поверхность (1500–2200 см 2 ). Новая кора занимает около 72 % всей площади коры и около 40 % массы головного мозга. В коре имеется 14 млрд нейронов, количество глиальных клеток примерно в 10 раз больше.
К основным бороздам коры относят боковую, центральную, теменно-затылочную, поясную и коллатеральную мозговые борозды. Они делят кору больших полушарий на доли. Различают шесть основных долей: лобную, теменную, височную, затылочную, краевую и островок. Каждая доля вторичными бороздами делится на извилины. По классификации К. Бродмана, кора разделена на 52 поля. С полями коры больших полушарий связаны ее регуляторные влияния на функции организма.
Кора головного мозга является наиболее молодой в филогенетическом отношении структурой мозга, осуществляющей у человека высшие интегративные функции контроля и регуляции соматических, висцеральных и психических функций.
В коре различают шесть горизонтальных слоев.
Выраженность отдельных слоев в различных областях коры не одинакова. По классификации К. Бродмана, основанной на цитоархитектонических признаках (формы и расположения нейронов), кора разделена на 52 поля. С полями коры больших полушарий связаны ее регуляторные влияния на функции организма (рис. 3.12).
Электрическая активность коры больших полушарий характеризуется специфическими ритмами определенной частоты и амплитуды, которые обозначаются термином электроэнцефалограмма (ЭЭГ).
Впервые у животных ЭЭГ была зарегистрирована в 1925 г. В. Правдич-Неминским, а у людей – психиатром Г. Бергером в 1929 г.
Для отведения ЭЭГ существуют два способа: биполярный и монополярный. При биполярном отведении регистрируется разность потенциалов между двумя активными электродами. При монополярном отведении регистрируется разность потенциалов между активной точкой на поверхности головы и индифферентной точкой головы или лица, где электрические процессы минимальные и их можно условно принять за нуль (обычно это сосцевидный отросток черепа или мочка уха). Выбор того или иного варианта записи зависит от целей исследования. В исследовательской практике шире используется монополярный вариант регистрации, поскольку он позволяет изучать изолированный вклад работы той или иной зоны мозга в изучаемый процесс.
Международная федерация обществ электроэнцефалографии приняла так называемую систему «10-20», позволяющую точно указывать расположение электродов. В соответствии с этой системой у каждого испытуемого точно измеряют расстояние между серединой переносицы и твердым костным бугорком на затылке, а также между левой и правой ушными ямками. Возможные точки расположения электродов разделены интервалами, составляющими 10 или 20 % этих расстояний на черепе. При этом для удобства регистрации весь череп разбит на области, обозначаемые буквами: F – лобная, О – затылочная, Р – теменная, Т – височная, С – область центральной борозды. Нечетные номера мест отведения относятся к левому, а четные – к правому полушарию. Символом Cz – обозначается отведение от верхушки черепа.
Вопрос о генезе ЭЭГ является довольно сложным, вместе с тем, исходя из современного уровня знаний, можно утверждать, что ЭЭГ является результатом суммации постсинаптических потенциалов корковых нейронов. Причиной возникновения синхронных постсинаптических изменений в корковых нейронах могут быть циклические таламокортикальные взаимодействия, в которых таламические нейроны играют роль своеобразных ритмоводителей, или пейсмекеров.
В настоящее время электрическая активность коры больших полушарий мозга детально изучена и анализ ее изменений с использованием разнообразных компьютерных программ широко используется в клинической практике.
В ЭЭГ человека выделяют следующие ритмы (рис. 3.13).
Выделяют также некоторые эквиваленты альфа-ритма, имеющие другую локализацию и чувствительные к другим видам модальности. В области роландовой борозды регистрируется мю-ритм (роландический), отвечающий блокадой на проприорецептивные раздражения. В височной коре регистрируется тау-ритм, который подавляется звуковыми стимулами. Каппа-ритм имеет также одну частотную характеристику с альфа-ритмом. Он регистрируется в височной области при подавлении альфа-ритма в других областях в процессе умственной деятельности.
Бета- ритм имеет частоту 18–30 Гц и амплитуду колебаний около 2–20 мкВ. Этот ритм регистрируется в прецентральной и фронтальной коре, а также и в других областях коры при различных видах интенсивной деятельности.
Тета-ритм включает колебания с частотой 4–8 Гц, амплитудой 5–100 мкВ и чаще наблюдается во фронтальных зонах коры, выражен в гиппокампе. Связан этот ритм с поисковым поведением, усиливается при эмоциональном напряжении.
Дельта-ритм состоит из самых медленных волн с частотой 0,5–4,0 Гц, амплитудой 20–200 мкВ. Возникает при естественном и наркотическом сне, а также при некоторых патологических состояниях коры.
В ответ на раздражение рецепторов, периферических нервов, ядер таламуса и других образований, связанных с проведением сенсорных сигналов, с поверхности коры регистрируются так называемые вызванные потенциалы. Они представляют собой последовательность из нескольких позитивных и негативных волн, которая длится в течение 300–400 мс после стимула.
Различают две группы вызванных потенциалов: первичные ответы (ПО) и вторичные ответы (ВО). Первичные ответы представляют собой двухфазные позитивно-негативные колебания, которые регистрируются с наименьшим латентным периодом (20–100 мс) в пределах проекционных зон коры при раздражениии специфических афферентов. Эта особенность первичных ответов была использована при исследовании топической организации коры больших полушарий и при идентификации связей между различными структурами мозга.
Предполагается, что позитивная фаза (направленная вниз) ПО обусловлена суммацией синаптических потенциалов тел и базальных дендритов пирамидных клеток. Негативная фаза (направленная вверх) связана с активацией апикальных дендритов этих же нейронов в поверхностных слоях коры.
Вторичные ответы (компоненты ВП в интервале 100–300 мс) регистрируются в ассоциативных корковых зонах, примыкающих к проекционным зонам. Вторичные ответы отражают процессы, связанные с передачей возбуждения от первичных проекционных зон к ассоциативным зонам и с рабочей настройкой корковых нейронов путем изменения их возбудимости.
Если испытуемому дается инструкция нажимать кнопку на второй из двух последовательных сигналов, то после ряда сочетаний первый сигнал (звуковой щелчок) начинает вызывать волну ожидания, которая продолжается до подачи второго сигнала (вспышки света). Волна ожидания представляет собой медленное негативное колебание с амплитудой порядка 40 мкВ, которая регистрируется от поверхности головы в зоне лобных и теменных долей. Предполагают, что негативная волна, предшествующая движению испытуемого, является электрофизиологическим коррелятором процесса подготовки к целенаправленному движению.
Функциональные зоны коры больших полушарий. В коре больших полушарий выделяют сенсорные, моторные и ассоциативные зоны.
В современной физиологии в двигательной коре выделяют первичную и вторичную моторные области.
Первичная двигательная область коры находится в области прецентральной извилины (поля 4 и 6), которая находится кпереди от центральной (Роландовой) борозды. На медиальной поверхности полушария, рядом с первичной областью обнаружена вторичная двигательная область.
Двигательная кора имеет соматотопический принцип организации, характеризующийся правильной пространственной проекцией соматических мышц. Зоны управления движениями кистей рук и мимической мускулатурой являются наиболее обширными и характеризуются самым низким порогом. Эти области помимо моторного выхода из коры имеют самостоятельные сенсорные входы от кожных и мышечных рецепторов. В этой связи они были названы первичной и вторичной мотосенсорной корой (Мс I и Mc II ).
В постцентральной извилине (поля 1, 2, 3) находится первичная соматосенсорная область. Сюда через специфические ядра таламуса приходят афферентные проекции от рецепторов кожи и двигательного аппарата. Эти проекции имеют соматотопическую организацию. В районе сильвиевой борозды, отделяющей височную долю от теменной и лобной, находится вторая соматосенсорная область. Эти области кроме афферентных входов содержат эфферентные выходы. Поэтому их называют первичной и вторичной сенсомоторными зонами (См I и См II ).
Первичная (17 поле) и вторичная (поля 18 и 19) зрительные области локализованы в затылочной доле. В правое полушарие проецируются рецепторы левой половины сетчатки, а в левое полушарие – правой половины сетчатки. В каждом полушарии происходит совмещение зрительных полей обоих глаз, что лежит в основе бинокулярного зрения. Электрическое раздражение зрительной коры может вызвать зрительные ощущения. Вторичная зрительная область контролирует зрительное внимание и управление движениями глаз.
Первичная проекционная зона слухового анализатора локализована в верхнем крае височной доли (поля 41 и 42). Для этой области характерно тонотопическое представительство кортиева органа. Повреждение коркового представительства слухового анализатора затрудняет восприятие и пространственную локализацию источника звука, а также распознание временных характеристик звука.
Рядом с проекционными зонами расположены ассоциативные поля, в состав которых входит ряд областей теменной, височной и лобной долей. Для нейронов ассоциативной коры свойственно отвечать на раздражение различных модальностей. Они участвуют в интеграции сенсорной информации и в обеспечении связей между чувствительной и двигательной корой, что является физиологической основой высших психических функций.
С помощью микроэлектродных методов исследования корковых нейронов в соматосенсорной коре кошки американскому исследователю В. Маунткаслу (1957 г.) удалось обнаружить факты, проливающие свет на принципы переработки сенсорной информации в неокортексе. Было обнаружено, что при погружении микроэлектродов перпендикулярно поверхности коры, все встречаемые клетки отвечали на раздражитель одной и той же модальности. При погружении же микроэлектродов под углом к поверхности коры встречаемые ими нейроны отвечали на раздражение различной модальности. На основании этих экспериментальных фактов В. Маунткасл пришел к заключению, что соматосенсорная кора организована в элементарные функциональные единицы (колонки), ориентированные перпендикулярно поверхности коры. Кортикальная колонка представляет собой обрабатывающее устройство с входом и выходом. Диаметр такой колонки порядка 500–1000 мкм определяется горизонтальным распространением терминалей афферентного таламокортикального волокна и вертикальной ориентацией дендритов пирамидных клеток.
Каждая функциональная колонка состоит из нескольких структурных микромодулей, объединяющих пять-шесть гнездообразно расположенных нейронов (рис. 3.15). В состав микромодуля входит несколько пирамидных клеток. Благодаря тесному сближению апикальных дендритов пирамидных клеток возможны электротонические связи, обеспечивающие синхронную работу микромодуля.
В состав микромодуля входят звездчатые клетки, с которыми контактируют таламокортикальные волокна. Некоторые звездчатые клетки с тормозной функцией имеют длинные аксоны, распространяющиеся в горизонтальном направлении.
Аксоны пирамидных клеток образуют возвратные коллатерали, которые могут обеспечивать облегчающие влияния в пределах микромодуля, и тормозные взаимодействия между микромодулями через интернейроны.
Работами Д. Хьюбела и Т. Вязеля был подтвержден принцип колончатой организации для зрительной коры. Здесь были обнаружены группы клеток (колонки), связанные с разной глазодоминантностью. Было замечено, что всякий раз, когда регистрирующий микроэлектрод входил в кору обезьяны перпендикулярно ее поверхности, он встречал клетки, лучше реагирующие на стимуляцию только одного глаза. Если же электрод вводили с наклоном и как можно более параллельно поверхности коры, то клетки с разной глазодоминантностью чередовались. Полная смена доминантного глаза происходила примерно через каждый 1 мм.
Открытие нового метода окраски нейронов с применением микроинъекций фермента пероксидазы хрена позволило проследить путь отдельных аксонов, приходящих из латеральных коленчатых тел (ЛКТ) в зрительную кору. Было установлено, что каждый аксон из ЛКТ проходит через нижние слои и оканчивается разветвлением в IV слое, принося информацию от одного глаза в одни участки. Показано, что каналы передачи зрительной информации от разных глаз в зрительной коре, так же, как и в ЛКТ таламуса, не пересекаются.
В коре присутствуют также клетки с бинокулярными свойствами, реагирующие на сигналы от обоих глаз. В верхних слоях коры при переходе от колонки одной глазодоминантности к другой существуют промежуточные зоны, где смена доминантности происходит не скачком, а постепенно, проходя через промежуточные стадии. В промежуточных зонах находятся нейроны, реагирующие на сигналы от обоих глаз.
В зрительной коре обнаружены также колонки, избирательно реагирующие на направление движения или на цвет.
Колончатый принцип организации нейронов присущ и другим областям коры.
Функциональная асимметрия полушарий мозга является важнейшим психофизиологическим свойством головного мозга человека. Еще в 1961 г. П. Брока, а в 1974 г. К. Вернике обнаружили локализацию центра речи в височной доле левого полушария.
В опытах на больных «с расщепленным мозгом» (перерезка мозолистого тела) были получены новые данные, проливающие свет на функциональную асимметрию полушарий. Оказалось, что по ряду психофизиологических функций доминирует не левое, а правое полушарие. В этой связи возникла концепция частичного доминирования полушарий.
При исследовании речи было показано, что словесный информационный канал контролируется левым полушарием, а несловесный канал (голос, интонация) – правым.
Хотя с речевыми функциями преимущественно связано левое полушарие, тем не менее правое полушарие также обладает языковыми функциями, а именно – может понимать письменную речь. С помощью правого полушария больные с рассеченным мозгом могут не только «читать» вопросы, но и «отвечать» на них левой рукой. Это связано с тем, что мышцы конечностей и туловища левой стороны тела контролируются моторной корой правого полушария и наоборот. Исключение составляют некоторые мышцы лица, которые контролируются обоими полушариями.
В сенсорной сфере функциональная асимметрия полушарий отчетливо проявляется при зрительном восприятии. Показано, что правое полушарие воспринимает зрительный образ целостно, сразу во всех подробностях, легче решает задачу различения предметов и опознания визуальных образов, которые трудно описать словами, создает предпосылки конкретно-чувственного мышления. Левое полушарие оценивает зрительный образ расчлененно, аналитически, при этом каждый признак (форма, величина и др.) анализируется раздельно.
В эмоциональной сфере правое полушарие обусловливает преимущественно древние, отрицательные эмоции, контролирует проявления сильных эмоций, в целом оно более «эмоционально». Левое полушарие связано в основном с положительными эмоциями, контролирует проявление более слабых эмоций.
Таким образом, знак эмоций зависит от соотношения активности левой (ЛФК) и правой (ПФК) фронтальной коры. Это правило В. Геллер представила в виде двух неравенств:
ЛФК > ПФК = положительные эмоции,
ПФК > ЛФК = отрицательные эмоции.
Абстрактное мышление и сознание связывают в основном с левым полушарием. При выработке условного рефлекса в начальной фазе доминирует правое полушарие, а во время упрочения рефлекса – левое.
Функциональную специализацию левого и правого полушарий головного мозга долгое время считали присущей только человеку, связывая ее с появлением речи. Однако латеральная специализация наблюдается уже у животных, обнаруживая сходство со специализацией полушарий человека.
Левое полушарие птиц, грызунов, хищников и приматов имеет преимущественное отношение к коммуникативным функциям, заученным формам поведения, тонким двигательным манипуляциям. Правое полушарие связано с эмоционально-окрашенным и зрительно-пространственным поведением. Левое полушарие важно для ориентировочно-исследовательской деятельности животного. В левом полушарии преобладают «когнитивные медиаторы» – ДА, АХ, ГАМК, а медиаторы, наиболее тесно связанные с мотивационно-эмоциональным поведением, – серотонин, норадреналин – в большем количестве находят в правом полушарии.
Физиология человека и животных
Разделы
14. Основы физиологии коры больших полушарий. Представление о кортикализации функций. Цитоархитектоника коры больших полушарий
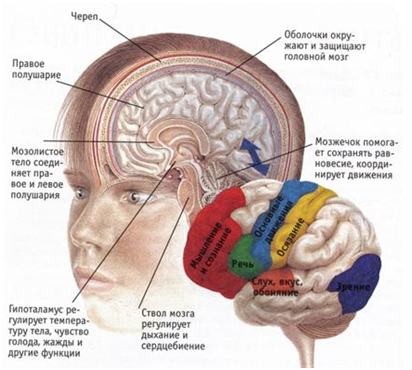
Каждое полушарие разделено бороздами на доли: лобную (здесь находятся вкусовая, обонятельная, двигательная и кожно-мускульная зоны), теменную (двигательная и кожно-мускульная зоны), височную (слуховая зона) и затылочную (зрительная зона). Каждое полушарие отвечает за противоположную ей сторону тела. В функциональном отношении полушария неравнозначны. Левое полушарие – «аналитическое», отвечает за абстрактное мышление, навыки письменной и устной речи. Правое полушарие – «синтетическое», отвечает за образное мышление.
 Колончатая организация зон коры. Кора больших полушарий состоит из сложных многоклеточных ансамблей, так называемых колонок (Маунткасл). Колонка является элементарным блоком сенсомоторной коры, где осуществляется переработка информации от рецепторов одной модальности. Колонка – это функциональное объединение нейронов, расположенных в «цилиндрике» (колонке) диаметром 0,5–1 мм, включающем все слои коры и содержащем несколько сотен нейронов. В эти мини-колонки входят:
Колончатая организация зон коры. Кора больших полушарий состоит из сложных многоклеточных ансамблей, так называемых колонок (Маунткасл). Колонка является элементарным блоком сенсомоторной коры, где осуществляется переработка информации от рецепторов одной модальности. Колонка – это функциональное объединение нейронов, расположенных в «цилиндрике» (колонке) диаметром 0,5–1 мм, включающем все слои коры и содержащем несколько сотен нейронов. В эти мини-колонки входят:
– нейроны, получающие входные сигналы от подкорковых структур (например, от ядер таламуса);
– нейроны, получающие входные сигналы от других областей коры;
– все нейроны локальных сетей, образующие вертикальные клеточные колонки;
– нейроны, передающие выходные сигналы от колонки назад к таламусу, другим областям коры, нейронам лимбической системы.
Соседние колонки могут частично перекрываться, а также взаимодействовать друг с другом по механизму латерального торможения и осуществлять саморегуляцию по типу возвратного торможения. Несколько простых вертикальных колонок могут с помощью межколоночных связей объединяться в более крупную единицу, перерабатывающую информации – модулярную колонку.
Кортиколизация функций – это возрастание в филогенезе роли коры больших полушарий в анализе и регуляции функций организма и подчинении себе нижележащих отделов ЦНС. Например, регуляция таких двигательных функций, как прыжки, ходьба, бег и выпрямительные рефлексы у низших позвоночных (амфибий) полностью обеспечивается стволом мозга. У кошек перезка ствола мозга между средним и промежуточным мозгом существенно не влияет на выпрямительные рефлексы, но нарушает локомоцию (бег, прыжки) на некоторое время. Выключение коры больших полушарий у обезьян и у человека приводит к потере не только локомоции, но и выпрямительных рефлексов.
Кора большого мозга делится на:
— древнюю, которая на ряду с другими функциями обеспечивает обоняние и соответствующее взаимодействие систем мозга;
— старую, включающую поясную извилину и гиппокамп;
— новую, особенно развита у человека, максимальна в передней центральной извилине.
Типы клеток коры:
1) Пирамидные клетки. Ориентированы вертикально по отношению к поверхности коры и и имеют тело треугольной формы. От тела вверх идет длинный Т-образно ветвящийся дендрит, а вниз от основания тела нейрона – аксон, идущий вниз из коры или в другие области коры. Дендриты покрыты мелкими шипиками (области синаптического контакта).
2) Звездчатые клетки. Они имеют короткие сильно ветвящиеся дендриты и аксоны, формирующие внутрикорковые связи.
3) Веретеновидные клетки. Имеют длинный аксон, который ориентирован либо вертикально, либо горизонтально.
В процессе филогенеза, т.е. в процессе развития более сложных и высокоорганизованных организмов происходит возрастание значения высших отделов ЦНС в жизни организма – так называемая кортиколизация функций, т.е. подчинение сложных реакций организма коре больших полушарий. Все, что приобретается организмом в течение индивидуальной жизни (онтогенеза) связано с функцией больших полушарий. Именно в них осуществляются функции, связанные с высшей нервной деятельностью.
Кора больших полушарий образует 6 слоев:
1 – первый слой, верхний, молекулярный, представлен в основном ветвлениями восходящих дендритов пирамидных нейронов, регулирующих уровень возбудимости коры больших полушарий;
2 – второй слой, наружный зернистый, состоит из звездчатых клеток, обеспечивает циркуляцию возбуждения в коре, имеет отношение к кратковременной памяти;
3 – третий слой, наружный пирамидный, обеспечивает корко-корковые связи различных извилин мозга;
4 – четвертый слой, внутренний зернистый, содержит звездчатые клетки, здесь заканчиваются специфические таламокортикальные пути, которые начинаются от рецепторов анализаторов;
6 – шестой слой – это слой полиморфных, или веретеновидных. клеток, образующих корково-таламические пути.
В слоях 1 и 4 происходит восприятие и обработка поступающей в КБП информации, Нейроны 2 и 3 слоев осуществляют кортикокортикальные ассоциативные связи. В слоях 5 и 6 преимущественно находятся нейроны, аксоны которых образуют эфферентные волокна, выносящие информацию из КБП.



