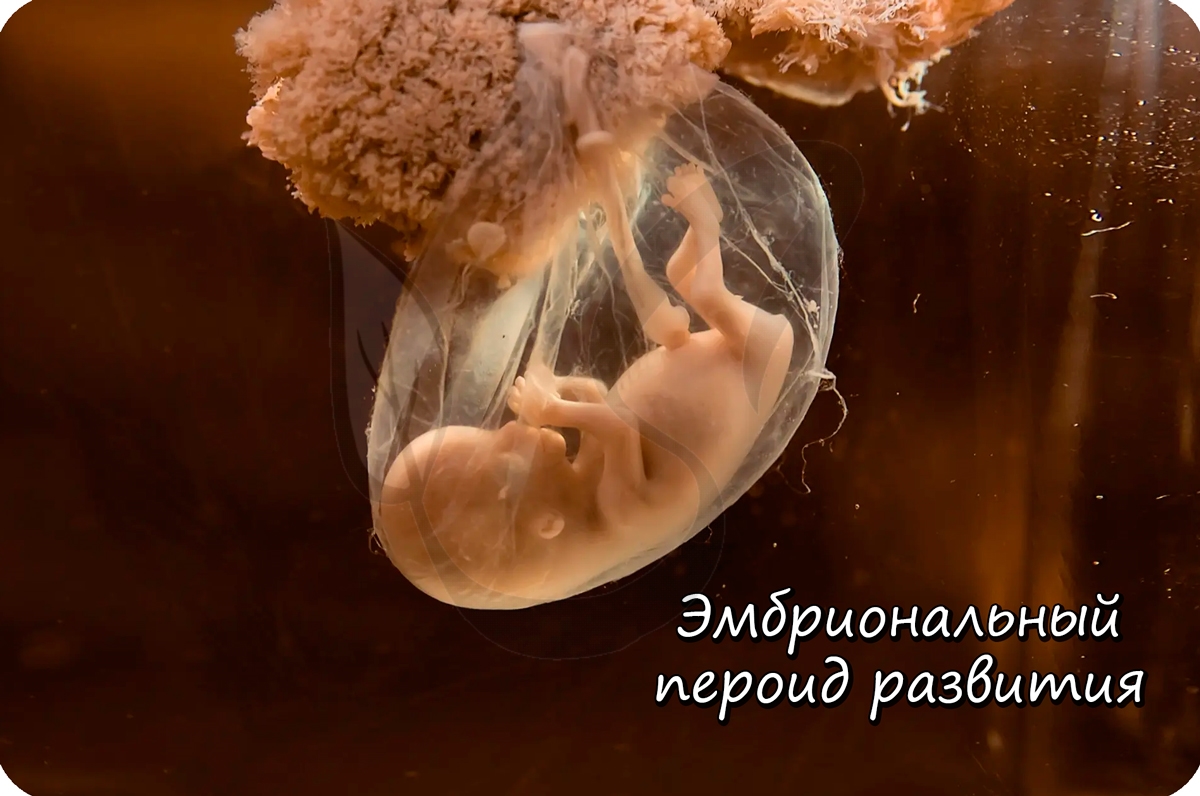
Эмбриональное развитие
От момента образования зиготы и до выхода зародыша из яйцевых оболочек длится эмбриональный период развития.
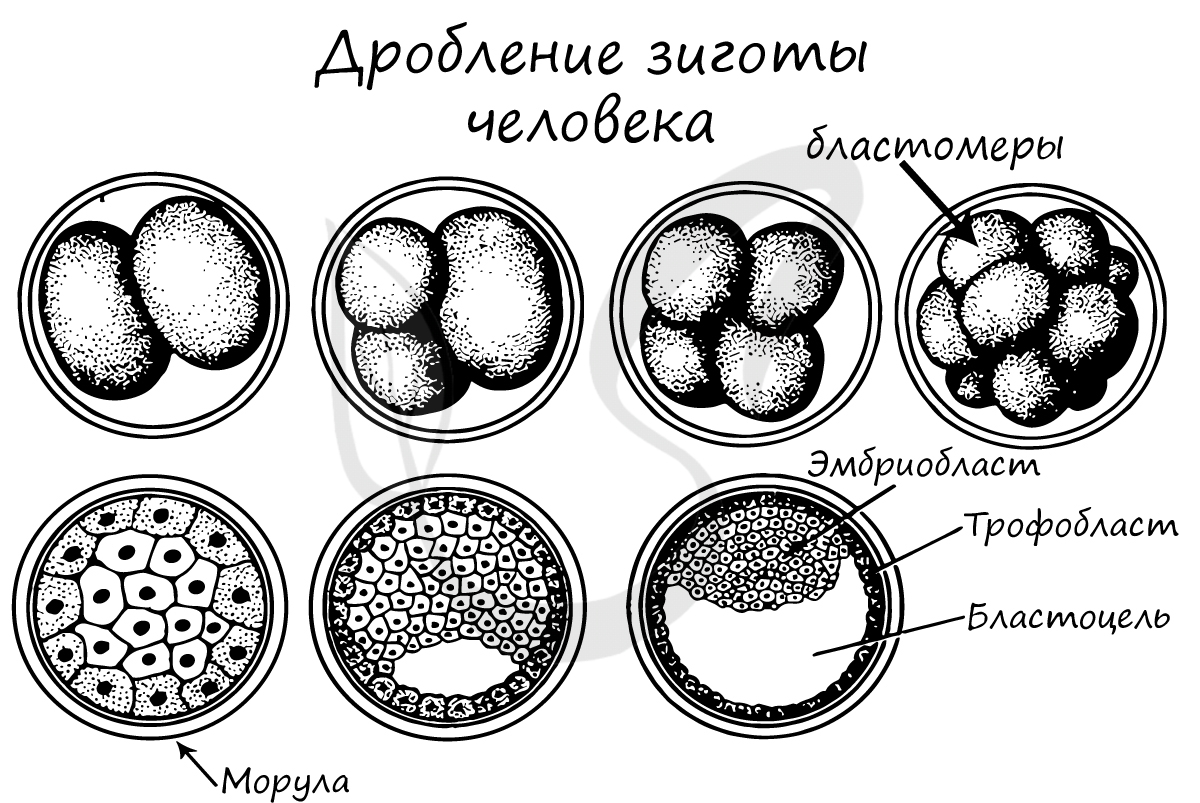
Дробление зиготы
Важная особенность дробления в том, что не происходит увеличение в размере зародыша: клетки дробятся (делятся) настолько быстро, что не успевают накопить цитоплазматическую массу. Дробление зиготы человека является полным неравномерным асинхронным.
Бластуляция

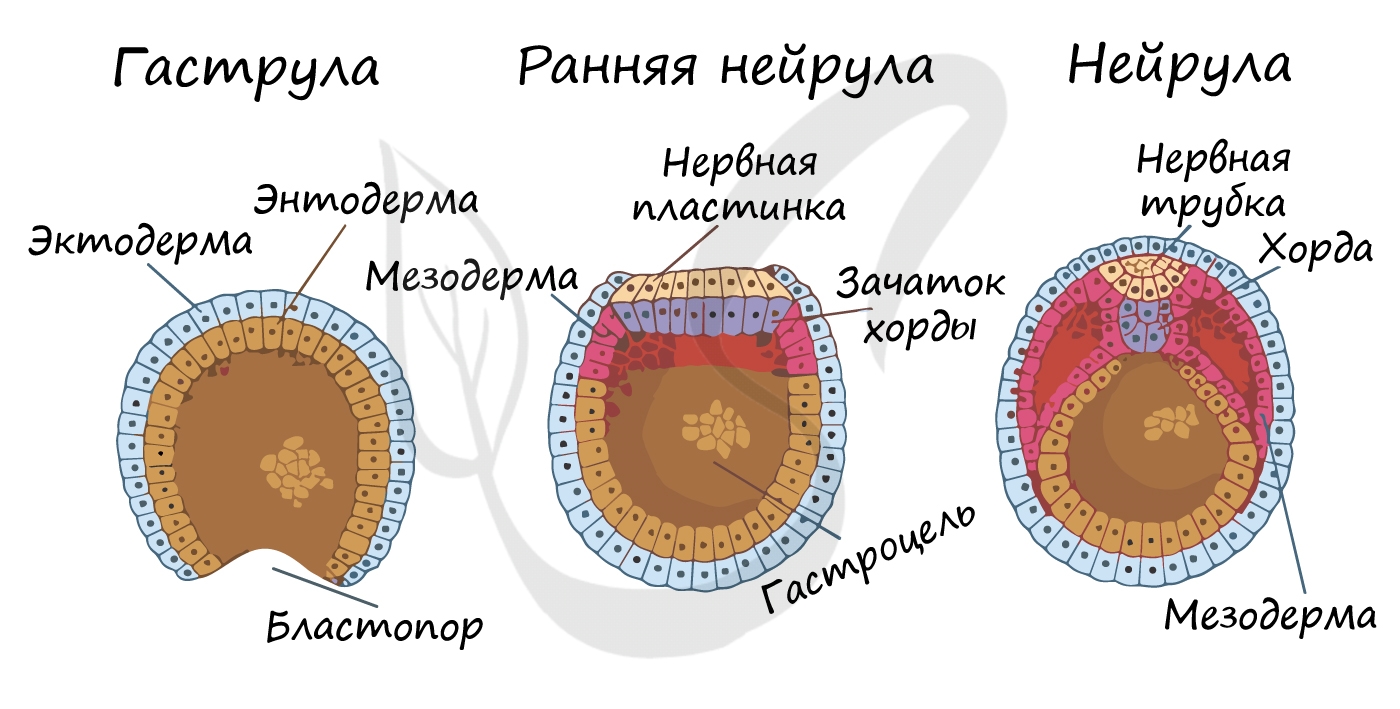
Гаструляция (греч. gaster — желудок, чрево)
Гаструляцией называют стадию эмбрионального развития, в ходе которой клетки, возникшие в результате дробления зиготы, формируют три зародышевых листка: эктодерму, мезодерму и энтодерму.

У первичноротых животных на месте первичного рта (бластопора) образуется ротовое отверстие. К первичноротым относятся: кишечнополостные, плоские, круглые и кольчатые черви, моллюски, членистоногие.
У вторичноротых на месте бластопора формируется анальное отверстие, а ротовое отверстие образуется на противоположном полюсе. К вторичноротым относят хордовых и иглокожих (морских звезд, морских ежей).


Нейрула
Эта стадия следует за гаструлой. Ранняя нейрула представляет собой трехслойный зародыш, состоящий из энто-, экто- и мезодермы. На этапе нейрулы происходит закладка отдельных органов.

Все три зародышевых листка требуют нашего особого внимания, а также понимания того, какие органы и структуры из них образуются.

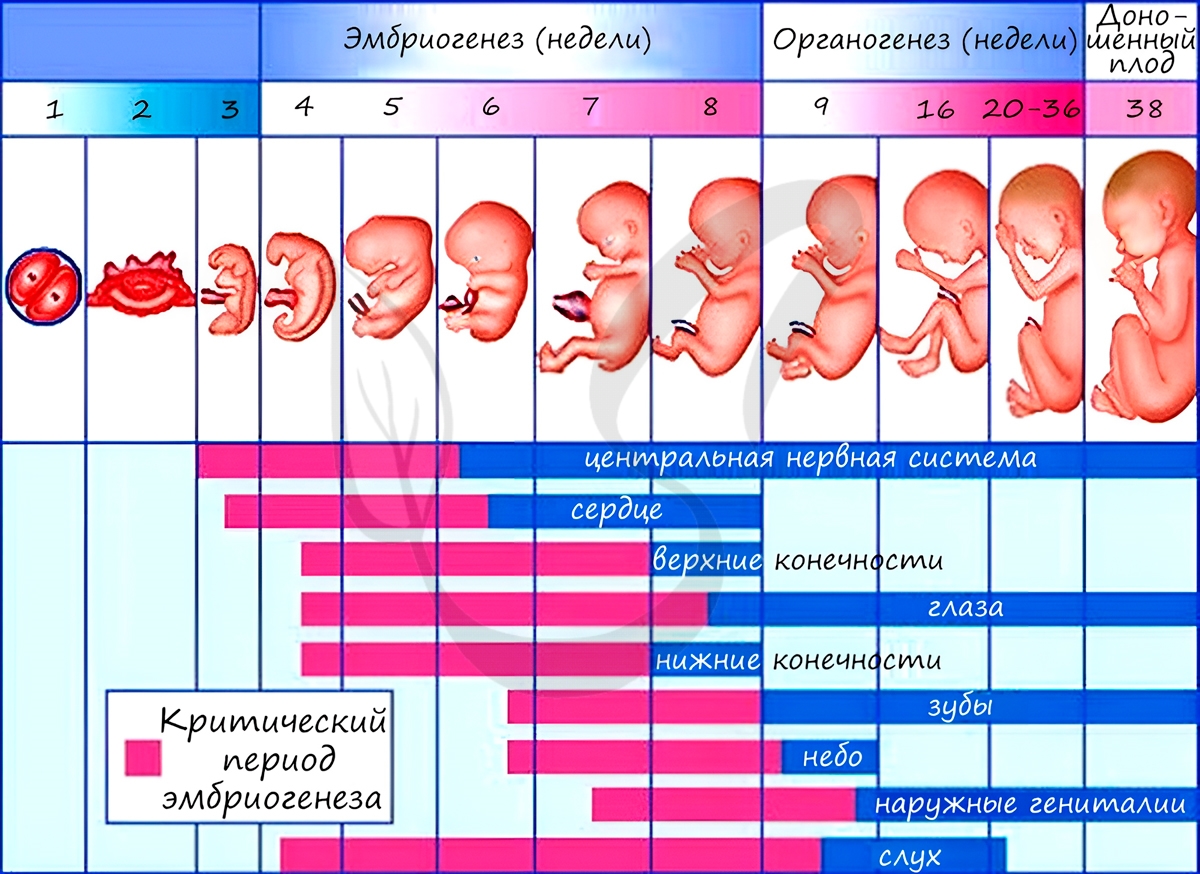
Из зародышевых листков образуются ткани, органы и системы органов. Такой процесс называется органогенезом. В период закладки органов важное значение имеет воздержание матери от вредных привычек (алкоголь, курение), которые могут нарушить процесс дифференцировки клеток и привести к тяжелейшим аномалиям, уродствам плода.
Некоторые лекарства также могут оказывать на плод тератогенный эффект (греч. τέρας — чудовище, урод), приводя к развитию уродств. Периоды закладки органов и система органов вследствие их большой важности носят название критических периодов эмбриогенеза.
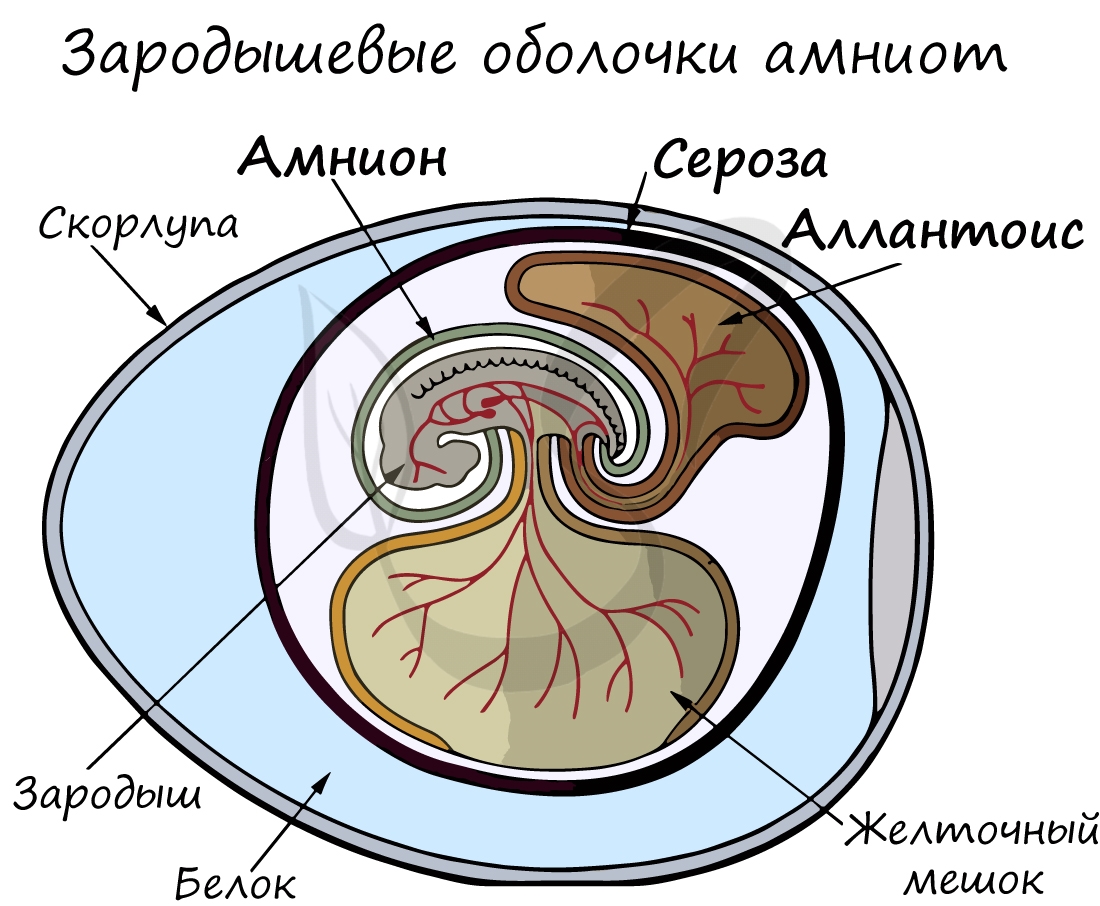
Анамнии и амниоты
К анамниям относятся рыбы, земноводные.

Зародышевый орган, аллантоис, является органом дыхания и выделения.
За счет особых оболочек, развивающихся в ходе эмбрионального развития, амниона и серозы, у амниот формируется амниотическая полость. В ней находится зародыш, окруженный околоплодными водами. Благодаря такому гениальному устройству, амниотам для размножения и развития более не нужно постоянное нахождение в водоеме, они «обрели независимость» от него.
© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Путешествие во времени: как судьба первых клеток эмбриона влияет на его дальнейшее развитие и риск заболеваний
коллаж автора на основе изображений с сайта depositphotos.com
Автор
Редакторы
19 марта 2021 года в научном журнале Science были опубликованы две научные статьи о самых ранних эмбриональных мутациях человека. 15 исследователей из американских Клиники Мэйо и Йельского университета под совместным руководством профессоров Алексея Абызова и Флоры Ваккарино разработали неинвазивный метод воссоздания клеточной родословной для каждого живущего человека и применили его для анализа иерархических деревьев клеточных клонов у двух живых людей и одного погибшего плода. Этот анализ помог приблизиться к моменту, когда оплодотворенная яйцеклетка разделилась на два первых бластомера, и сделать несколько важных выводов. Мы расспросили Алексея Абызова о том, что нового им удалось узнать.
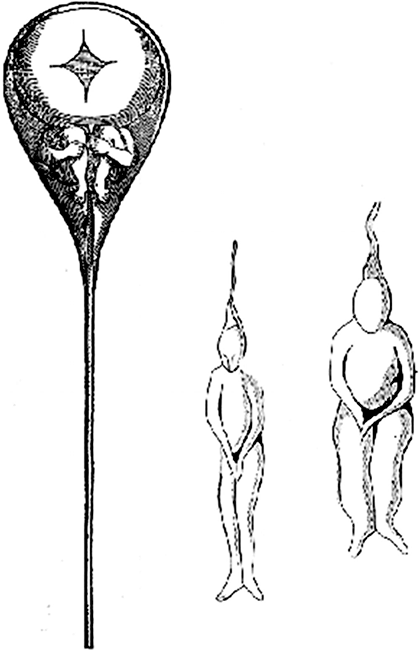
Рисунок 1. Так, по мнению преформистов, выглядел маленький гомункул в мужских сперматозоидах. Рисунок голландского математика и физика Николаса Хартсокера, 1695 г.
Несколько столетий назад в биологии господствовало учение о преформизме. Лучшие умы человечества ничего не знали о генах, а потому считали, что в половых клетках изначально находится гомункул — маленький человечек (рис. 1). И тут мнения сразу разделились. Овисты поместили гомункул в ооцит, а анималькулисты — в сперматозоид, отведя женским половым клеткам скромную роль питательной среды [1].
По мере развития эмбриологии идеи преформизма были признаны ошибочными. В XIX веке российский естествоиспытатель Карл Бэр (рис. 2) установил, что зародыши человека на ранних этапах развития похожи вовсе не на гомункулов, а на червячков и рыбок. С открытием генов стало понятно, что в половых клетках содержится не зачаток готового организма, а «рецепт» по его «приготовлению», записанный в ДНК. Но это породило массу новых вопросов. Как этот «рецепт» определяет судьбу разных клеток? Какие факторы могут вмешиваться, помимо генетического кода? Могут ли особенности развития организма, склонности к тем или иным заболеваниям закладываться уже на уровне самых первых бластомеров?

Рисунок 2. Карл Бэр — российский естествоиспытатель, сформулировавший законы зародышевого сходства («законы Бэра»).
Этим темам посвящено немало научных исследований. Но судьба первых бластомеров и ее влияние на дальнейшее формирование организма по-прежнему во многом остаются terra incognita.
Человек асимметричный
Человек относится к живым организмам с двусторонней симметрией. Однако в реальности наше тело далеко не так симметрично, как может показаться на первый взгляд. В первую очередь это касается некоторых органов: все знают, что сердце и желудок находятся слева, а печень справа. Многие парные органы тоже неодинаковы: правое легкое у людей состоит из трех долей, а левое — всего из двух, верхний полюс правой почки обычно находится на уровне 12 межреберья, а левой — на уровне 11 ребра. У мужчин левое яичко зачастую находится ниже правого, а размеры его меньше [2], [3]. У женщин немного различаются размеры правой и левой молочных желез. Правое и левое полушария головного мозга выглядят одинаково, но работают немного по-разному, различается объем правой и левой лобных и затылочных областей [4].
Некоторые заболевания тоже «выбирают» определенную половину тела. Например, так происходит с двигательными симптомами при болезни Паркинсона, при гемимегалоэнцефалии (патология, при которой сильно увеличивается одно из полушарий головного мозга, и в нем происходят патологические изменения) [5]. Исследования в некоторых популяциях показали, что у женщин рак молочной железы чаще возникает слева, но при правосторонней локализации зачастую хуже прогноз и раньше возникают метастазы [6], [7].
Можно привести еще много примеров асимметрии в теле человека, связанных как с нормой, так и с патологией. Это очень интересная сфера для научных исследований, и зачастую она уводит на клеточный уровень, заставляя разбираться в изменениях, которые происходили в клетках, начиная с самых ранних этапов эмбриогенеза.
Формирование организма происходит иерархично, в нем прослеживается несколько уровней клеточной организации. Еще до прикрепления к стенке матки зародыш разделяется на две части. Трофобласт (рис. 3) представляет собой пузырек диаметром 0,1 мм и является предшественником плаценты. Внутри него находится эмбриобласт — скопление клеток, из которого затем образуется эмбрион. Постепенно эмбриобласт принимает форму диска — становится так называемым зародышевым диском. Он ориентируется в пространстве, в нем выделяются передняя и задняя, правая и левая части, появляются три зародышевых листка — эктодерма, энтодерма и мезодерма (рис. 4), — дающие начало разным тканям [8]. Каждая клетка занимает отведенное ей место и передает своим потомкам инструкцию о том, что делать дальше. Конечно же, интересно отследить эти процессы, разобраться в «родословной» каждой клетки в организме человека.
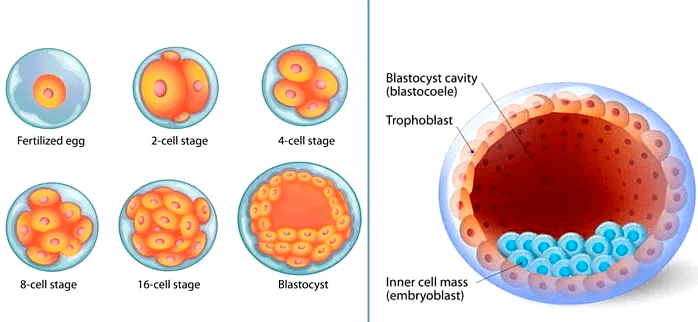
Рисунок 3. Самые ранние стадии эмбриогенеза. На изображении справа трофобласт выглядит как полая сфера. Внутри, у одного из ее полюсов, находится скопление клеток голубого цвета — это эмбриобласт.
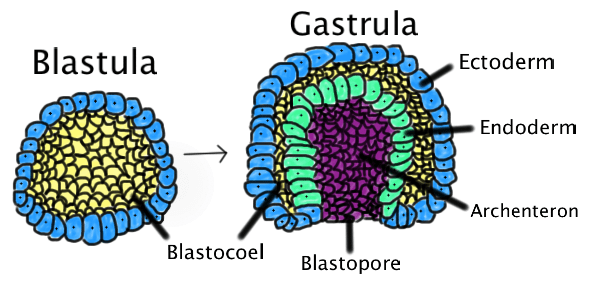
Рисунок 4. Три зародышевых листка: энтодерма, эктодерма и мезодерма. Из эктодермы образуются наружные покровные ткани и нервные структуры; из мезодермы — мышцы, органы кровеносной, выделительной и половой систем; из энтодермы — пищеварительная система и другие внутренние органы.
Пытаясь исследовать развитие эмбрионов человека, ученые сталкиваются с массой сложностей, как технического, так и этического характера. Эксперименты над людьми и эмбрионами проводить запрещено, поэтому приходится исследовать клеточные линии ретроспективно. Постфактум это можно сделать, только если остаются определенные «следы», мáркеры. К счастью, такие маркеры есть. Это соматические мутации.
Маяки для путешествия в прошлое
Мутагенез — двигатель эволюции, и он постоянно происходит во всех живых клетках. Наследственные мутации присутствуют в ооцитах и сперматозоидах и передаются от родителей детям. Они обнаруживаются во всех клетках тел детей. Соматические мутации возникают в соматических клетках, из которых состоит тело человека, и которые не участвуют в половом размножении. Они присутствуют не во всем теле, а только в потомках той клетки, в которой изначально произошло изменение. Например, пигментный невус (родинка) представляет собой скопление таких клеток. К распространенным мутациям в пигментных невусах относится мутация V600E в гене BRAF — он кодирует фермент киназу, способствующую пролиферации (размножению) клеток (эта мутация встречается и в клетках злокачественной опухоли — меланомы).
В настоящее время широко применяется термин «послезиготные (постзиготические) мутации» — они появляются после слияния половых клеток и начинают возникать буквально с первых минут эмбриогенеза. Исследования показывают, что на каждое из пяти первых делений после оплодотворения яйцеклетки приходится по 1–2 новых соматических мутации, а в дальнейшем этот показатель увеличивается [9]. Большинство из этих мутаций — нейтральные: они не приносят ни вреда, ни пользы. Но некоторые влияют на важные процессы в клетках и, накапливаясь, со временем приводят к заболеваниям. Например, в 2018 году было установлено, что у здоровых людей старшего возраста на одну клетку слизистой оболочки пищевода приходится от 20 до более чем 2000 мутаций, и некоторые из них связаны с онкологическими заболеваниями [10].
В первую очередь мутации изучают для того, чтобы разобраться в последствиях, к которым они приводят в организме — например, вызывают те или иные заболевания или повышают предрасположенность к их развитию. Но также ученые придумали, как с помощью анализа изменений в генах отслеживать клеточные линии. Если одна и та же соматическая мутация встречается во многих клетках, то можно с высокой степенью достоверности утверждать, что у этих клеток был общий предок. Изучив генетические изменения в разных клетках и тканях организма с помощью технологий секвенирования [11], можно судить о времени возникновения этих изменений — вплоть до первых делений зиготы (оплодотворенной яйцеклетки с диплоидным набором хромосом). Тут работает принцип: чем больше клеток являются носителями мутации, тем у более ранней клетки-предшественницы она возникла, и наоборот. Это дает возможность выстраивать иерархические деревья (рис. 5, 6), которые помогают ученым понять, как события на самых ранних этапах эмбриогенеза влияют на дальнейшее развитие организма, и какой вклад вносят разные клеточные клоны в формирование тканей.
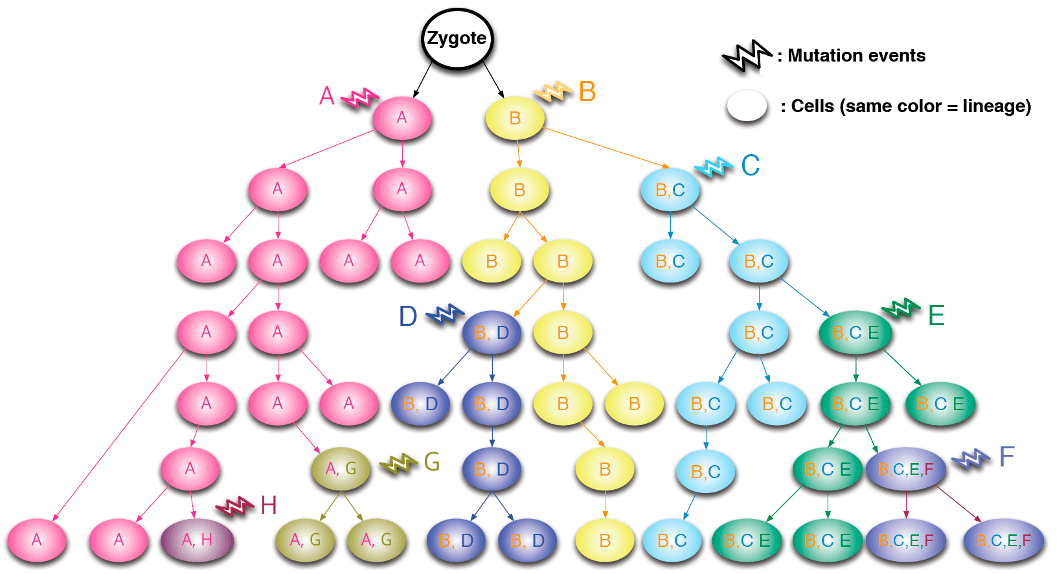
Рисунок 5. Как можно отследить родословную клеточных клонов, изучая соматические мутации? Разными буквами и цветами обозначены новые мутации, которые возникают в материнских клетках и передаются их потомкам. Видно, как изменения в генах накапливаются с каждым новым поколением.
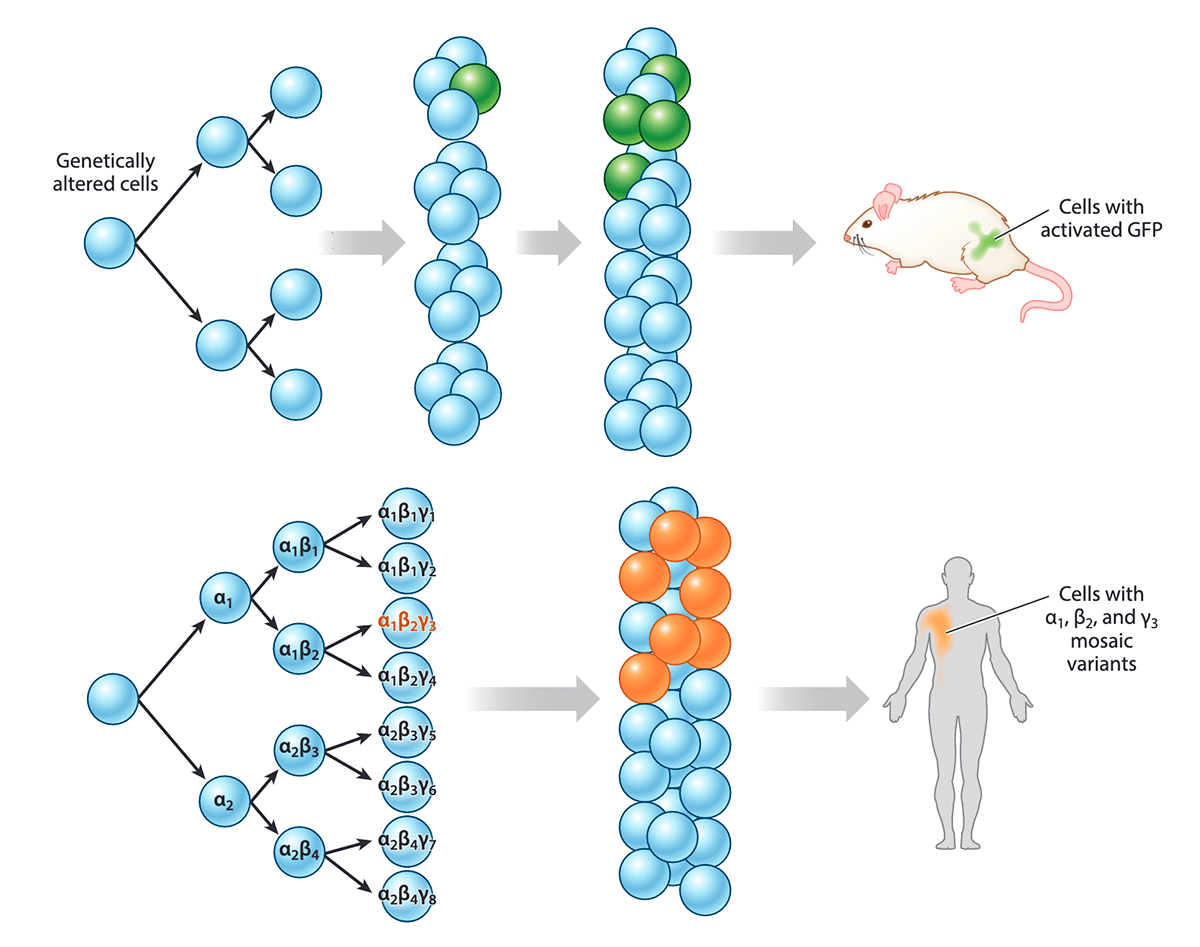
Рисунок 6. Соматические мутации и мозаицизм. Голубым цветом обозначены клетки без мутаций. Видно, как в одной из клеток возникает новая мутация. У мыши клетка с изменением в генах (активация GFP — зеленого флуоресцентного белка) отмечена зеленым цветом — ее создали искусственно с помощью генной инженерии. Ниже представлен пример похожего (но возникшего естественным путем) мозаицизма у человека: клетки с мутациями обозначены оранжевым цветом. Например, большое оранжевое пятно на теле человека на рисунке может быть огромным врожденным невусом. А может быть (и чаще всего так бывает), эта мутация нейтральная и никак себя не проявляет — ее можно обнаружить только с помощью генетического анализа.
Ранее уже проводили исследования родословных клеток с помощью анализа послезиготных мутаций. Например, удалось оценить скорость мутагенеза на ранних стадиях развития эмбриона, установить, что два первых бластомера вносят неодинаковый вклад в развитие тела человека — асимметрия возникает уже после первого дробления зиготы. Авторы одной научной работы, опубликованной в 2017 году, указывают соотношение вклада ранних эмбриональных клеток в формирование популяции клеток крови 2:1 [10]. Соотношение потомков разных клеточных клонов может различаться в разных тканях, оно коррелирует с пространственной организацией органов. Ученые продолжают разбираться в причинах и закономерностях этих процессов. Возможно, это поможет не только лучше изучить ранние этапы эмбриогенеза, но и разработать новые методы профилактики, диагностики, лечения некоторых заболеваний.
Что нового удалось выяснить?
Ученые под руководством Алексея Абызова (рис. 7) использовали для реконструкции событий, происходящих на ранних этапах эмбриогенеза, клетки, полученные от двух живых людей (здоровая 66-летняя женщина и 29-летний мужчина, страдающий синдромом Туретта [12]) и погибшего плода. У живых людей взяли образцы кожи рук и ног, выделили фибробласты и получили из них несколько десятков клонов искусственных стволовых клеток, а затем секвенировали их ДНК. Также для исследования были взяты образцы мочи, крови и слюны. У погибшего плода использовали клетки головного мозга, фибробласты твердой мозговой оболочки, ткань селезенки [8], [13].

Рисунок 7а. Алексей Абызов — один из руководителей исследования. Алексей — выпускник МФТИ; д. ф.-м. н.; с 2014 года руководит научной лабораторией в Клинике Мэйо; ассоциированный профессор в медицинском колледже при этой клинике; приглашенный профессор в Университете Миннесоты и Йельском университете. Ранее мы уже публиковали обзоры исследований, которыми руководил Алексей: «Геномная головоломка: открой в себе мозаика» [14] и «Как искусственные модели головного мозга и омиксные технологии пригодятся в борьбе с аутизмом» [15]. А еще Алексей участвует в качестве спикера и ментора в школах Future Biotech (летние школы в 2012 и 2013 годах, зимняя школа в 2020 г.).

Рисунок 7б. Лаборатория Алексея Абызова в клинике Мэйо
То, как из одной зиготы образуется 37 триллионов клеток, из которых состоит человеческое тело, является одной из величайших загадок биологии. Мы разработали метод воссоздания личной клеточной родословной в самом раннем, послезиготном развитии по клеткам живущего человека, которые могут быть неинвазивно собраны и проанализированы в любом возрасте. Описанная методология обладает потенциалом к дальнейшему развитию, но уже сейчас может применяться для воссоздания клеточной родословной на более поздних стадиях развития, а также в других многоклеточных организмах помимо человека.
Изучение соматических мутаций в ДНК образцов тканей и реконструкция событий, происходивших на ранних этапах эмбриогенеза, показали, что первые бластомеры вносят неравномерный вклад в формирование тканей тела. Уже после первого дробления яйцеклетки выделяется доминантная клеточная линия, представленная в тканях на 70–90%, и рецессивная, которой достается только 10–30%. Самая большая разница была обнаружена в крови — 90:10. В моче соотношение доминантного и рецессивного клонов составило 70:30 — 80:20, а в слюне оно было либо как в крови, либо в промежутке между показателями крови и мочи.
Приставка метил- означает, что произошла реакция метилирования, и к молекуле прикрепилась метильная группа. Если это происходит с основаниями ДНК, то последовательность гена не меняется, а вот его работа может измениться, и довольно сильно. Такие трансформации относятся к сфере эпигенетики. Кстати, на «Биомолекуле» есть статья «Неканоническая ДНК» [17], из которой читатель, ориентирующийся в «языке» генетического кода, может узнать, чем на самом деле отличается цитозин от 5-метилцитозина, а еще — чем тимин отличается от урацила. Рекомендуем также обратиться к статье «Молекулы и эпигеном» [18] из спецпроекта по эпигенетике.
Отсюда напрашивается предположение: возможно, рецессивный клон потому и становится рецессивным, что в его клетках хуже восстанавливается ДНК, и им на это требуется больше времени. Есть и альтернативная гипотеза: возможно, рецессивный клон предназначен в первую очередь для формирования трофобласта и плаценты. Разобраться поможет изучение ткани плаценты — это тема для будущих научных работ. Вот как об этом говорит сам Алексей:
Главным открытием оказалось то, что ветви в воссозданных генеалогических древах оказались разнопредставленными, то есть асимметричными, по количеству клеток в органах каждого человека. В частности, одна из двух клеток после первого дробления зиготы является прародителем абсолютного большинства (70–90%) клеток в каждом исследуемом человеке. Возможным объяснением может быть то, что эта клетка предназначена в первую очередь для формирования организма взрослого человека, в то время как вторая клетка предназначена для создания плаценты, которая будет питать растущий эмбрион. Исследователи также высказали альтернативную гипотезу, что разнопредставленность может быть результатом разной скорости роста клеток и/или разной способности справляться с повреждением ДНК.
Изучая ткани погибшего плода, ученые обнаружили, что клеточные клоны в некоторых областях мозга упорядоченно распределяются вдоль переднезадней оси. Такой характер распределения говорит, что оно предопределяется событиями, которые происходят в клетках-прародительницах этих клонов. Судьба их потомков запрограммирована с самого начала.
Еще одно интересное наблюдение: ранние мутации нередко в точности совпадают с геномными вариациями, которые встречаются в популяции в целом. Иными словами, одни и те же изменения в генах независимо друг от друга, случайным образом, происходят у разных людей. Это наталкивает на мысль, что в геноме есть области, особенно чувствительные к мутагенезу. Именно в них чаще всего и происходят «поломки» [8], [13].
Это исследование стало важным шагом на пути изучения событий раннего эмбриогенеза. Ученые разработали неинвазивный метод воссоздания клеточной родословной для каждого живущего человека, и в будущем его можно использовать для анализа родословных клеток на разных стадиях развития у разных многоклеточных организмов. Помимо фибробластов кожи, крови, слюны и мочи, можно взять любой другой биоматериал, полученный от живого человека: например, стул, мазки из половых путей женщины, мужскую семенную жидкость. Это открывает возможности для дальнейших исследований.
Выявить болезнь до того, как она началась
В исследовании приняли участие два пациента с миелопролиферативными заболеваниями (редким типом злокачественных заболеваний крови) в возрасте 63 и 34 лет. Ученые провели у них биопсию красного костного мозга, секвенировали геном здоровых и злокачественных клеток, а затем составили их родословную. Исследователи пытались выяснить, когда у этих больных возникла мутация JAK2, вызвавшая заболевание. Оказалось, что у 63-летнего больного это произошло примерно в 19 лет, а у 34-летнего — в 9 лет [19].
О мозаицизме, его связи с заболеваниями и посвященной этой теме научной работе, которую ранее опубликовала группа Алексея Абызова, мы подробно рассказывали в статье «Геномная головоломка: открой в себе мозаика» [14].
В другом исследовании 2012 года оказалось, что клональный мозаицизм у людей с сóлидными опухолями встречается чаще, чем у здоровых, причем связь оказалась сильнее, когда материал для анализа брали до постановки диагноза и начала лечения. Клональный мозаицизм был более распространен среди людей, у которых анализ ДНК проводили за год до того, как им диагностировали лейкемию [21].
Эти исследования показывают, что первые мутации, приводящие к онкологическим и другим заболеваниям, могут возникать задолго до того, как появляются симптомы, изменения в лабораторных анализах или патологические изменения, которые можно рассмотреть на снимках.
Выявлять болезни по генетическим анализам задолго до того, как появятся первые симптомы — заманчивая перспектива. В 2012 году была опубликована научная работа, авторы которой описали первый успешный опыт применения персональной геномики. Ученые проанализировали у одного пациента геном и массив других молекулярных данных и смогли выявить у него сахарный диабет второго типа на очень ранней стадии. Самое интересное, что этим пациентом стал руководитель исследования. Мы рассказывали об этой работе в статье «Упреки в нарциссомике» [22].
А могут ли некоторые из этих мутаций возникать еще во время внутриутробного развития? Играют ли роль в их возникновении события, происходящие на ранних этапах эмбриогенеза? И может ли быть так, что из-за «слабости» генома клетки из рецессивного клона становятся «уязвимыми точками» для развития ряда заболеваний, и их можно идентифицировать заранее? Это очень сложные вопросы, и вероятно, в будущем ученые найдут ответы и на них.
Эмбриональное развитие нервной системы позвоночных
Раннее развитие
Авторы
Нервная система всех позвоночных, включая человека, развивается из элементов наружного зародышевого листка – эктодермы. Этот процесс имеет определенные особенности у представителей разных групп, однако ему свойственны и общие для всех позвоночных закономерности.
В период гаструляции у высших позвоночных (у человека это конец 1-й недели развития и совпадает с имплантацией в стенку матки) происходят активные перемещения клеточного материала зародыша. В первой фазе гаструляции образуются два эмбриональных зародышевых листка – эпибласт (верхний листок) и гипобласт (нижний). Клетки эпибласта постепенно расходятся, образуя заполненную жидкостью амниотическую полость. Во второй фазе гаструляции небольшая группа клеток эпибласта, сформировавшая в дне амниотической полости зародышевый щиток, образует первичную полоску и гензеновский узелок. Последующая миграция клеток этих структур вглубь зародыша приводит к формированию среднего листка зародыша – мезодермы. Гаструляция завершается у всех позвоночных образованием трех зародышевых листков: эктодермы, мезодермы и энтодермы, а также формированием осевого комплекса зачатков органов. Особое значение на этом этапе развития принадлежит т.н. головному отростку (нотохорду), формирующемуся из мигрирующих клеток гензеновского узелка. К концу гаструляции формируются и все основные, соответствующие разным группам животных провизорные органы (желточный мешок, амнион, аллантоис, хорион, плацента), выполняющие защитные и питательные функции для эмбриона. Их число в ходе эволюции увеличивается. У человека гаструляция завершается к третьей недели внутриутробного развития.
Головной отросток дает начало развитию нотохорды – оси будущего зародыша. Клетки нотохорды и формирующейся затем хорды оказывают индуцирующее влияние на дифференцировку прилежащего к ним участка эктодермы в нервную пластинку и далее в нервную трубку (рис. 1). Как только развивается нотохорд, расположенная над ним эктодерма начинает утолщаться и формирует нервную пластинку, элементы которой интенсивно размножаются и дифференцируются, превращаясь в узкие цилиндрические нейроэпителиальные клетки, отличные от соседних клеток покровного эпителия. Основной причиной формирования нервной пластинки и замыкания ее в нервную трубку является преобразование нейроэпителиальных клеток, связанное с изменением ориентации компонентов их актинового цитоскелета. В результате интенсивного деления и неравномерного роста нейроэпителия происходит его инвагинация с последующим формированием нервной трубки.
Эмбриональное развитие ЦНС у млекопитающих и человека обычно разделяют на: эмбриональный (первые 6 недель), фетальный (с 6 до 24 недели) и перинатальный периоды (с 24 недели до рождения) (табл. 1).
| Стадия развития | Возраст плода (недели) | Основные морфологические изменения в развитии мозга |
| Эмбриональный период | ||
| Формирование и разделение герминативного слоя | 2 | Нейрональная пластинка |
| Дорсальная индукция: первичная нейруляция | 3–4 | Формирование: нервной трубки, нервного гребня и ее производных; закрытие рострального и каудального нейропоров; парных крыловидных пластинок |
| Вентральная индукция: теленцефализация | 4–6 | Развитие конечного мозга и структур лица; формирование мозговых пузырей; развитие оптических и обонятельных плакод; появление зачатков ромбовидного мозга и мозжечка |
| Фетальный период | ||
| Нейрональная и глиальная пролиферация | 6–16 | Пролиферация клеток в вентрикулярной и субвентрикулярной зонах формирующихся отделов мозга, включая неокортекс; ранняя дифференциация нейробластов и глиобластов; процессы апоптоза; миграции клеток в стенке мозга |
| Миграция | 12–24 | Миграция нейронов в формирующиеся отделы мозга; формирование мозолистого тела полушарий и других проекционных путей ЦНС |
| Перинатальный период | ||
| Регионализация | 24 – до рождения | Завершение процессов миграции и формирование основных отделов мозга; синаптогенез; созревание популяций нейронов и глиальных клеток |
| Миелинизация | 24 – до 2х лет после рождения | Окончательное созревание морфологической структуры мозга; миелинизация основных трактов и связей; активное функциональное развитие важнейших отделов головного мозга (особенно ассоциативных областей полушарий) |
Содержание

Рис. 1. Схема ранней стадии развития зародыша человека (формирование головного отростка – нотохорды).
1 – головной отросток; 2 – эктодерма, 3 – первичная полоска, 4 – первичная ямка, 5 – энтодерма, 6 – формирующийся аллантоис, 7 – прехордальная пластинка, 8 – полость желточного мешка, 9 – амниотическая полость (по: Данилов, Боровая, 2016, с изменениями)

Рис. 3. Первичная индукция формирования нейроэпителия будущей нервной пластинки (по: Development of Nervous System, 2006, с изменениями).
Chd – хордин, Ng – ноггин, IMZ – эмбриональная мезодерма, BPM4 – костный морфогенетический белок, TGFr – рецептор к фактору роста опухолей

Рис. 4. Участие ряда сигнальных молекул, транскрипционных факторов и ростовых факторов в формировании нервной пластинки и нервной трубки на ранних этапах эмбриогенеза нервной системы (по: Development of Nervous System, 2006, с изменениями).
СH – хордин, NG – ноггин, FS – фоллистатин, Shh – sonic hedgehog, BMP – костный морфогенетический белок, FGF – фактор роста фибробластов, TGF – фактор роста опухолей, Pax3, 4, 6 – транскрипционные факторы, S – мезодермальные сомиты, NC – нервный гребень
Эмбриональный период (1–23 стадии по Carnegie)
В этот период развития на дорсальной стороне зародыша происходит обособление особого участка нейроэпителия и формирование нервной пластинки (neuronal plate) и начинаются процессы нейруляции.
На стадии нейруляции происходит формирование нескольких важных структур нервной системы: образуется нервная пластинка с последующим образованием нервной трубки и нервного гребня (рис. 2). Нейруляция у человека начинается в конце 3-й недели и полностью завершается к концу 4-й недели.
Вскоре после образования нервной пластинки (приблизительно на 18-е сутки у человека) она прогибается вдоль продольной оси, ее края приподнимаются и формируются нервный желобок и нервные валики. Позднее края нервных валиков смыкаются по срединной линии и образуется замкнутая нервная трубка. Краниальный и каудальный участки нервной трубки долго остаются незамкнутыми, их называют соответственно передним и задним нейропорами. Передний нейропор закрывается на 23–26-й день развития, а задний – на 26–30-й день.

Процесс нейрональной индукции связан с синтезом ряда биологически активных соединений, которые действуют на формирование нервной пластинки и нервной трубки. На клетки первичной эктодермы действует большое количество сигнальных молекул, индуцирующих процесс образования нейроэпителия и нейрональных стволовых клеток, из которых будут формироваться все элементы нервной ткани. Среди этих факторов необходимо отметить хордин (chordin), ноггин (noggin) и фоллистатин (follistatin), синтезируемые клетками первичной мезодермы, образующей нотохорду (будущая хорда и позвоночник). Они блокируют действие другого морфо-генетического фактора – BMP (bone morphogenetic protein), синтезируемого клетками эктодермы и индуцируют их дифференцировку в направлении образования нейроэпителия нервной пластинки (рис. 3).
Уже на ранних этапах развития зародыша нервная трубка на значительном протяжении разделяется проходящей по вентрикулярной поверхности пограничной бороздой, sulcus limitans, на два отдела: дорсальный – крыловидную пластинку, и вентральный – базальную пластинку. Участки мозга, развивающиеся из крыловидной пластинки, содержат ассоциативные и сенсорные ядра, из базальной – моторные и вегетативные. Самая ростральная часть (prosencephalon) не содержит базальной пластинки и целиком происходит из крыловидной. Отделы головного мозга, содержащие производные обеих пластинок – средний, задний, продолговатый – часто объединяют названием «ствол мозга».
На этапе формирования нервных желобков дифференцировку вентральной части нервной трубки (базальной пластинки) и развитие мотонейронов оказывает регулирующее влияние фактор Shh (sonic hedgehog) секретируемый сначала нотохордой, а затем хордой и вентральной частью самой нервной трубки. Дорсальную часть нервной трубки (крыловидную пластинку) контролируют морфогенетические белки BMP4 и MBP7, секретируемые клетками эктодермы, и ряд других ростовых и транскрипционных факторов: Pax 3, 4, 6 – транскрипционные факторы, FGF8 – фактор роста фибробластов, GDNF – нейротрофический фактор глии, BDNF, NT3,4 – нейротрофические факторы мозга и др. (рис. 4).
Нервная трубка на ранних стадиях эмбриогенеза представляет собой многорядный нейроэпителий, состоящий из нейроэпителиальных клеток. В дальнейшем в нервной трубке дифференцируется 4 концентрических зоны:
Вентрикулярная (VZ) зона состоит из делящихся клеток цилиндрической формы. Вентрикулярные или иначе матричные клетки являются по сути нейрональными стволовыми клетками, т.е. предшественниками нейронов и клеток макроглии. Субвентрикулярная зона (SVZ) состоит из клеток, сохраняющих высокую пролиферативную активность и являющихся потомками матричных клеток. Промежуточная (плащевая или мантийная) зона (PZ) состоит из клеток, переместившихся из вентрикулярной и субвентрикулярной зон – это зона активной миграции и дифференцировки молодых нейронов (нейробластов) и глии (глиобластов). Наружная (маргинальная MZ) зона содержит нервные волокна и отростки находящихся ниже нейронов. Нейробласты достигают мест своего окончательного расположения в структурах ЦНС; утрачивают способность к делению и в дальнейшем дифференцируются в зрелые нейроны. Глиобласты продолжают делиться и дают начало новым популяциям глиальных клеток: астроцитам и олигодендроцитам.
Образование нервного гребня
После смыкания валиков и образования нервной трубки, на ее боковых поверхностях выселяется группа клеток, формирующих т. н. нервный гребень (neural crest) (рис. 2). Клетки нервного гребня активно и целенаправленно мигрируют на большие расстояния в зародыше и способны дифференцироваться в разнообразные зрелые элементы тканей и органов. Миграция клеток определяется не только и не столько наличием свободного межклеточного пространства и отсутствием механических преград на пути перемещения, сколько взаимодействием мигрирующих клеток с молекулами межклеточного матрикса (коллаген, ламилин, фибронектин, аминоглюкозгликаны и др.). Формирование фенотипа клеток нервного гребня определяют многочисленные ростовые и дифференцирующие факторы, действующие на клетки гребня по ходу их миграции. В качестве примера можно привести процесс дифференцировки клеток туловищного отдела нервного гребня в нейроны симпатических ганглиев или в клетки хромаффинной ткани мозгового вещества надпочечников (рис. 5).

Клетки нервного гребня образуются почти на всём протяжении замыкающейся нервной трубки. Клетки из различных участков нервного гребня дифференцируются не одинаково. Разная дифференцировка клеток наблюдается как по длине гребня, так и по глубине залегания в нем. Из гребня могут образовываться и нервные узлы и большая часть структур черепа. Уникальность этой структуры позволила современным ученым даже считать нервный гребень четвертым зародышевым листком, наряду с эктодермой, энтодермой и мезодермой. Вот неполный список производных клеток нервного гребня:
Фетальный период развития (24–46 стадии по Carnegie)
В этот период развития происходят наиболее значительные события в развитии мозга. Сразу после завершения нейруляции и образования первичных мозговых пузырей, в стенке эмбрионального мозга начинаются интенсивные процессы пролиферации и дифференцировки. Процессы пролиферации и дифференцировки захватывают широкую полосу клеток, расположенных между наружной (базальной) и внутренней (апикальной) поверхностями стенки мозговых пузырей. Они представляют собой нейрональные стволовые клетки (НСК) и развиваются из нейроэпителия нервной пластинки. НСК активно делятся и в процессе прохождения клеточного цикла претерпевают сложные превращения, связанные с последовательными перемещениями в нервной трубке. Перемещение осуществляются путем смешения ядросодержащих отделов клеток внутри формирующихся отростков. Этот процесс получил название интеркинетической ядерной миграции. Ядросодержащие тела клеток двигаются к поверхности нервной трубки, вблизи которой они остаются на некоторое время. Затем ядросодержащие отделы клеток опять перемещаются к вентрикулярной поверхности, после чего НСК втягивают свои отростки и вступают в очередной митотический цикл (M). В результате формируется одно из первых структурных образований развивающейся стенки нервной трубки – вентрикулярный слой (рис. 6а, б).
В настоящее время показано, что популяции клеток, составляющих вентрикулярный и формирующийся несколько позднее субвентрикулярный слой, неоднородна. Не все клетки, перемещающиеся в пределах стенки мозга во время митотического цикла, вступают в митоз у вентрикулярной поверхности. В зависимости от присутствия у клеток отростков и характера их контакта с поверхностями стенки мозга выделяют три класса клеток предшественников: монополярные, биполярные и неполярные (рис. 6в).
Биполярные клетки (или апикальные предшественники АР) представляют собой либо НСК клетки, либо клетки т. н. радиальной глии (RG), в которые НСК превращаются на самых ранних этапах нейрогенеза. Отличительной особенностью этих клеток является наличие отростков, контактирующих с апикальной и базальной поверхностями стенки мозга на всем протяжении клеточного цикла. Интеркинетические перемещения ядра происходят по этим отросткам и заканчиваются митозом у апикальной поверхности. Монополярные предшественники появляются на более поздних стадиях, когда в стенке мозга формируется субвентрикулярный слой, содержащий также как и вентрикулярный слой НСК. Ядра этих клеток претерпевают интеркинетические перемещения по цитоплазме отростков клеток предшественников, однако в процессе митотического цикла их апикальные или базальные отростки могут терять связь соответственно с апикальной или базальной поверхностями стенки мозга. Митозы происходят как в вентрикулярном слое, так и в верхней области субвентрикулярного слоя. Во внутренних слоях субвентрикулярной зоны у человека недавно были обнаружены клетки предшественники с неполярной морфологией. Характерной чертой этих клеток является ретракция отростков перед митозом и потеря их контакта с апикальной и базальной поверхностью стенки мозга. Они получили наименование «базальные предшественники (ВР).
Фактически мы имеем дело с двумя путями образования нейронов в развивающемся мозге. Это – путь прямого нейрогенеза, когда источником нейробластов являются непосредственно НСК и нейрогенная радиальная глия, т. е. апикальные предшественники с моно- или биполярной морфологией, и путь непрямого нейрогенеза, когда источником нейробластов служат промежуточные нейрональные предшественники, являющиеся потомками клеток радиальной глии, т. е. базальные предшественники. Непрямой путь кортикогенеза может выступать в роли быстрого увеличения количества нейронов в условиях ограниченного времени (каждое асимметричное деление радиальной глии через стадию промежуточного нейронального предшественника может давать два – четыре нейрона) и тем самым регулировать площадь и толщину стенки мозга.
Таким образом, на первых этапах формирования нервной системы, в стенке эмбрионального мозга формируется широкий слой пролиферирующих нейрональных предшественников разного типа, активность которых в дальнейшем приводит к формированию будущих популяций нервных и глиальных клеток в различных отделах мозга.

Рис. 6. Классические схемы перемещения клеток в вентрикулярном слое нервной трубки (а, б) и современное представление (в) о гетерогенности нейрональных предшественников. (по: Нейроонтогенез, 1985; Обухов, 2008, Pernavelas et al., 2002, с изменениями)
VZ, ISVZ,OSVZ, MZ – вентрикулярный, внутренний и наружный субвентрикулярный и маргинальный слои стенки мозга; apical, basal – апикальная (внутренняя) и базальная (наружная) поверхности мозговой стенки; Tc, Ts, G1, G2, M – стадии митотического цикла; I – период интерфазы; NB – нейробласт

Рис. 8. Схема развития мозговых пузырей (A) и формирования головной части зародыша позвоночных (Б) (по: Обухов, Андреева, 2017; Developmental neurobiology, 2005).
А: сомитомеры и мозговые пузыри костистых рыб и амниот (а), хрящевых рыб и амфибий (б). 1-11 – сомитомеры (будущие туловищные сомиты); 12 – закладка ушной капсулы; 13 – спинальный ганглий; 14, 15 – дорсальный (14) и вентральный (15) корешки спинномозгового нерва; 16 – полость целома; 17 – гипобранхиальная мускулатура; 18 – обонятельный орган, 19 – глазной пузырь; 20, 21 – жаберные щели (20) и дуги (21); III–X, XII – черепно-мозговые нервы

Рис. 10. Раннее развитие головного мозга и зоны экспрессии транскрипционных факторов, контролирующих формирование основных отделов головного мозга млекопитающих, вид сверху (А) и сбоку (Б) (Echevarria et al., 2003, с изменениями).
Pros, Tel, Die, Mes, Rhomb, Met, Myel – отделы головного мозга, Р1–Р6 – прозомеры, r1–r2 – ромбомеры, ANR – anterior neural ridge (передний мозговой организатор), ZLI – zona limitans interthalamica organizer (таламический организатор), ISO – isthmic organizer (организатор перешейка), SC, IC – верхние и нижние бугорки четверохолмия, P1–5 – прозомеры, r1–2 – ромбомеры; Pax 3/7, Ent, Otx, Foxg, Gbx – транскрипционные факторы и место их действия; стрелки – направление диффузии регулирующих факторов
Дифференцировка нервной трубки и формирование основных отделов мозга
Замыкание нервной трубки начинается в середине зародыша, затем процесс распространяется к головному и хвостовому концам эмбриона, где некоторое время остаются незамкнутыми отверстия – передний и задний нейропоры (рис. 7).

Рис. 7. Ранние этапы формирования нервной трубки на примере развития мозга человека (по: Nieuwenhuys R. et al., 1999).
A–D – реконструкция вида человеческого зародыша и начальных этапов формирования нервной трубки, E–H – поперечные срезы эмбриона на данных стадиях развития; 1 – эктодерма, 2 – нервная пластинка, 3 – отверстие амниона, 4 – мозговая пластинка, 5 – нервная складка, 6 – нервный желобок, 7 – нервная трубка, 8 – зачаток головного мозга, 9 – передний нейропор, 10 – задний нейропор, 11 – нервный гребень, 12 – крыловидная пластинка, 13 – латеральная пластинка, 14 – базальная пластинка, 15 – полость первичных мозговых желудочков, 16 – зачатки спинальных ганглиев
Еще на стадии замыкания нейропоров начинается ростро-каудальная дифференцировка нервной трубки зародыша. Нервная трубка (как полагают, под индуцирующим воздействием хорды) постепенно погружается в мезодерму зародыша и под влиянием мезодермальных сомитов разделяется на сегментарные участки – нейромеры или прозомеры. Сомиты располагаются по сторонам нервной пластинки и вдавливаются в нее, определяя конфигурацию будущих отделов мозга (рис. 8).
В дальнейшем головные сомиты сливаются и образуют три основных сегмента: премандибулярный, мандибулярный и гиоидный. Границей головных сегментов служит область ушной капсулы, за которой формируются от 2–3 до 10–12 туловищных сегментов (в зависимости от группы позвоночных). Параллельно формируется система черепно-мозговых нервов. Каждый сегмент иннервируется определенными парами нервов: премандибулярный – терминальным и глазодвигательным нервом (III); мандибулярный – тройничным (V) и блоковым (IV) нервами; гиоидный – отводящим (VI) и лицевым (VII) нервами. Следующие за головными два сегмента иннервируются соответственно языкоглоточным (IX) и блуждающим (X) нервами. Ростральные туловищные сомиты у высших позвоночных иннервируются системой добавочного нерва (XI), включающего в себя разное количество корешков в зависимости от числа туловищных сомитов. Подъязычный нерв (XII), иннервирующий гипобранхиальную мускулатуру, которая развивается из закладки туловищных сегментов, по своей функции аналогичен вентральным (соматомоторным) корешкам спинномозговых нервов, иннервирующих поперечнополосатую мускулатуру туловища и конечностей.
Передний конец трубки в конце 3-й недели развития из-за активных процессов пролиферации и миграции нейронов в стенке мозга расширяется и формирует 3 первичные мозговые пузыря. Лежащий краниально пузырь образует первичный передний мозг, Prosencephalon, средний пузырь – первичный средний мозг Mesencephalon, а из третьего пузыря развивается первичный задний мозг Rhombencephalon. Далее располагаются структуры формирующегося спинного мозга – Medulla spinalis (рис. 9).

Рис. 9. Развитие мозга человека (по: Шаде, Форд, 1976).
А–Б – стадии трех (а) и пяти (б–д) мозговых пузырей; вид сверху (А) и сбоку (Б); 1–3 – первичные: передний (1), средний (2), ромбовидный (№) мозг; 4 – закладка спинного мозга, 5 – глазной бокал, 6–10 – отделы мозга: конечный (6), промежуточный (7), средний (8), задний (9), продолговатый (10); 11 – полушария конечного мозга, 20 – мозжечок, 22 – спинной мозг, V–IX – черепно-мозговые нервы. Стрелки – изгибы нервной трубки (с. и. – среднемозговой, ш. и. – шейный, м. и. – мостовой)
Спинной мозг образуется из каудальных отделов нервной трубки. Он представляет собой часть ЦНС, в структуре которой наиболее отчетливо сохраняются черты эмбриональных стадий развития мозга позвоночных: трубчатый характер строения и сегментарность.
После формирования мозговых пузырей в нервной системе начинаются сложные процессы внутренней дифференцировки и роста. Уже на ранних этапах развития зародыша нервная трубка на значительном протяжении разделяется проходящей по вентрикулярной поверхности пограничной бороздой, sulcus limitans, на два отдела: дорсальный – крыловидную пластинку, и вентральный – базальную пластинку. Участки мозга, развивающиеся из крыловидной пластинки, содержат сенсорные ядра, из базальной – моторные и вегетативные. Ростральная часть нервной трубки не содержит базальной пластинки и целиком происходит из крыловидной. Отделы головного мозга, содержащие производные обеих пластинок – средний, задний, продолговатый – часто объединяют названием «ствол мозга».
Изменения в развитии нервной трубки сопровождаются образованием нескольких изгибов на границах закладки различных отделов мозга. В течение первых двух месяцев эмбрионального развития образуется основной (среднемозговой) изгиб, когда передний и промежуточный мозг загибаются вперед и вниз. Затем формируется еще два (шейный и мостовой) изгиба. Одновременно первый и третий первичные мозговые пузыри разделяются каждый на два. Наступает стадия пяти мозговых пузырей. Самым ростральным становиться конечный мозг (Telencephalon), затем – промежуточный (Diencephalon). За промежуточным идет средний мозг (Mesencephalon). Первичный задний мозговой пузырь разделяется на задний мозг (Metencephalon) и продолговатый мозг (Medulla oblongata). Прозенцефалон включает производные первых шести прозомеров (нейромеров) P1–P6. Из структур Р1 в дальнейшем формируется средний мозг. Прозомеры Р2 и Р3 развиваются соответственно в таламус и преталамус. Из прозомеров Р4–Р6 развивается конечный мозг и гипоталамус. Из более каудальных сегментов нервной трубки (ромбомеров) развиваются структуры ствола и спинного мозга.
После формирования мозговых пузырей (5–10 недели развития) в структурах формирующейся нервной системы происходят сложные процессы внутренней дифференцировки и роста различных отделов головного и спинного мозга.
Формирование отделов мозга находится под контролем т. н. «вторичных организаторов» – групп клеток, синтезирующих ряд морфогенетических факторов, градиент концентрации которых определяет направление миграции и дифференцировки разных структур мозга (табл. 2; рис. 9, 10).
| Ген | Место экспрессии | Функция |
| Dlx 1, Dlx 2, Dlx 5 | Субпаллиум (ганглионарные возвышения), промежуточный мозг | Миграция субпаллиальных нейробрастов, миграция нейронов в кору из ганглионарных возвышений переднего мозгового пузыря |
| Emx 1, Emx 2 | Конечный мозг | Пролиферация клеток в развивающемся мозге, миграция нейробластов |
| Lhx 1, Lhx 2, Lhx 5 | Передний мозг, кора полушарий | Формирование подкорковых и корковых (архикортекс) отделов полушарий |
| Nkx 2,1 Nkx 2,2 | Вентральные отделы полушарий | Пролиферация и миграция нейробластов в стриатуме |
| Otx 1, Otx 2 | Передний мозг, средний мозг, передние отделы ствола мозга | Формирование структуры полушарий, включая кору мозга |
| Pax 3, Pax 6 | Передний мозг | Миграция нейробластов в дорсальных отделах полушарий |
Развитие зачатка переднего мозга контролирует небольшая группа клеток, расположенная на верхушке нервной трубки и названная передним мозговым организатором (ANR – anterior neural ridge) и клетки на границе второго мозгового пузыря – zona limitans interthalamica (ZLI). Структуры среднего, заднего, продолговатого мозга и верхние сегменты спинного мозга контролируются еще одним организатором – isthmic organizer (ISO).
В перинатальный период заканчивается формирование внутренней структуры мозга. Начинается активная миелинизация головного и спинного мозга. Однако эти процессы не заканчиваются с рождением. Показано, что достаточно долго (месяцы и годы) после рождения происходит созревание и дифференцировка нервных структур и проводящих трактов. Более того, в настоящий период стало ясно, что во взрослый период происходит образование новых популяций нейронов и глиальных клеток за счет сохранения в мозге популяций НСК в структурах головного мозга.



