Параграф 72. о синтезе нуклеотидов
Автор текста – Анисимова Е.С.
Авторские права защищены. Продавать текст нельзя. Курсив не зубрить.
Замечания можно присылать по почте: exam_bch@mail.ru
https://vk.com/bch_5
ПАРАГРАФ 72. См. п.71, 54.
«О синтезе нуклеотидов».
Для чего нужен синтез нуклеотидов?
Синтез оснований и нуклеотидов нужен в основном
для синтеза из них ДНК и РНК.
АТФ обязательно нужен клетке для жизни в качестве формы энергии.
Другие функции нуклеотидов – в п. 70.
Если синтез нуклеотидов в клетке снижается:
(из-за плохого питания, снижения ПФП и т.д.),
то это приводит к снижению синтеза ДНК и РНК и далее –
к нарушению функций клетки, к старению и гибели клетки и всего организма.
О последствиях нарушения синтеза пуриновых нуклеотидов путем регенерации см. далее.
При синтезе нуклеотидов происходит синтез и соединение
трех частей нуклеотида:
азотистого основания, рибозы и фосфата.
Рибозо-5-фосфат образуется из глюкозы в ПФП.
(Если активность ПФП снижается,
то это приводит к снижению синтеза нуклеотидов, РНК и ДНК.
Причинами снижения ПФП могут быть гипогликемия, дефицит витаминов РР, В1,
низкая активность ферментов ПФП из-за плохого питания, дефектов их генов).
Для превращения Р-5-Ф в нуклеотид остается присоединить к нему основание.
Присоединение азотистого основания к рибозо-5-фосфату:
Способы присоединения оснований к Р-5-Ф разные:
при синтезе пиримидиновых нуклеотидов к Р-5-Ф
присоединяется готовое пиримидиновое основание,
которое называется ОРОТАТОМ.
При синтезе пуриновых нуклеотидов
к Р-5-Ф присоединяются (последовательно) несколько групп атомов,
постепенно образуя основание, которое называется ГИПОКСАНТИНОМ.
Р-5-Ф вступает в реакции после присоединения к нему двух фосфатов (пирофосфата);
соединение Р-5-Ф и пирофосфата называется фосфорибозил-пирофосфатом ФФПФ
и считается активной формой Р-5-Ф
(а реакция получения активной формы – активацией Р-5-Ф).
72. 1. Синтез п и р и м и д и н о в ы х нуклеотидов.
Первый образующийся пиримидиновый нуклеотид состоит:
из азотистого основания оротата, рибозы и фосфата.
Соединение оротата и рибозы (нуклеоЗид) называется оротидином,
соединение оротидина с фосфатом (нуклеоТид) называется ОротидинМоноФосфатом,
сокращенно – ОМФ.
Оротат синтезируется отдельно от Р-5-Ф,
а затем оротат присоединяется к рибозо-5-фосфату, образуя ОМФ.
Этапы синтеза пиримидиновых нуклеотидов:
1. Синтез карб/амоил/фосфата (КАФ) из Глн (его амидного азота) и СО2.
2. Синтез (основания) ОРОТАТА из КАФ и аспартата;
оротат отличается от урацила тем, что
в 6-м положении оротата есть СООН группа (которая впоследствии отщепляется).
3. Оротат присоединяется к Р-5-Ф, образуя нуклеоТид ОМФ.
4. От ОМФ отщепляется СО2,
в результате чего образуется УМФ;
реакция называется декарбоксилированием ОМФ,
катализируется ферментом ОМФ\декарбоксилазой
с участием кофермента пиридоксаль/фосфата (форма В6) – п.63.
5. Присоединение двух фосфатов (фосфорилирование)
превращает УМФ в УТФ;
источник двух фосфатов – 2 молекулы АТФ,
катализируют реакции (нуклеотид)киназы.
7. УМФ превращается в ТТФ
путем присоединения (см. формулы) СН3 группы, двух фосфатов и превращения рибозы в дезоксирибозу; источник СН3 группы – [СН3]-ТГФ.
72. 2. Синтез п у р и н о в ы х нуклеотидов.
Первый пуриновый нуклеотид, образующийся при синтезе пуриновых нуклеотидов,
состоит из гипоксантина, рибозы и фосфата.
Соединение гипоксантина и рибозы (нуклеоЗид) называется ИНОЗИНОМ,
соединение инозина с фосфатом (нуклеотТид) называется ИнозинМоноФосфатом,
сокращенно – ИМФ.
Гипоксантин не синтезируется отдельно от Р-5-Ф,
он синтезируется путем присоединения групп к Р-5-Ф.
Источниками групп при синтезе ИМФ являются:
3 аминокислоты: глутамин (отдает амидный азот), глицин, аспартат,
и [С1]-ТГФ и СО2.
Этапы синтеза пуриновых нуклеотидов:
2. ИМФ превращается в обычные нуклеотиды АМФ или ГМФ.
(Понятно при сравнении формул ИМФ и АМФ, гипоксантина и аденина:)
3. Превращение ИМФ в АМФ
происходит при замене ОН группы ИМФ (гипоксантина в составе ИМФ)
на NH2 группу;
источником NH2 группы является Аспартат,
требуется расщепление ГТФ в качестве источника энергии.
(См. формулы гипоксантина и гуанина, ИМФ и ГМФ:)
4. Превращение ИМФ в ГМФ
происходит при присоединении NH2 группы к ИМФ (к гипоксантину в составе ИМФ); источником NH2 группы является Глутамин, требуется расщепление АТФ в качестве источника энергии.
5. АМФ и ГМФ превращаются в АТФ и ГТФ
при присоединении фосфатов (при фосфорилировании),
источник фосфатов – АТФ,
катализируют реакции (нуклеотид)киназы.
Но при превращении АДФ в АТФ происходит СФ или ОФ – см. п. 23.
Этот путь синтеза нуклеотидов (из аминокислот) называется de novo («заново»)
и протекает в основном в печени.
В клетках крови и головного мозга
синтез пуриновых нуклеотидов путем de novo отсутствует,
поэтому они получают пуриновые нуклеотиды из печени (зависят от поставок пуриновых нуклеотидов из печени).
Когда пуриновые нуклеотиды разрушаются до оснований (аденин, гуанин, гипоксантин),
то основания можно не превращать в мочевую кислоту (п.72),
а использовать их для синтеза новых нуклеотидов –
достаточно присоединить к ним Р-5-Ф: далее.
72. 3. Р е г е н е р а ц и я пуриновых нуклеотидов.
Аденин (аденозин), гуанин и гипоксантин,
образующиеся при распаде нуклеотидов (см. п. 71),
могут не только превращаться в ураты,
но и использоваться для синтеза нуклеотидов.
Для превращения оснований в нуклеотиды
достаточно добавить к ним Р-5-Ф
(к аденозину нужно присоединить фосфат – получится АМФ).
Этот путь синтеза нуклеотидов из оснований
регенерацией пуриновых нуклеотидов.
Таким образом, пуриновые нуклеотиды могут синтезироваться не только из аминокислот
(т.е. путем de novo),
но и из продуктов распада нуклеотидов:
из нуклеозидов и азотистых оснований.
Значение регенерации пуриновых нуклеотидов:
она позволяет:
1) получить новые нуклеотиды для синтеза НК
(что особенно важно для клеток крови и мозга
из-за отсутствия в них синтеза путём de novo),
2) избежать синтеза избытка мочевой кислоты
из оснований, не потраченных на регенерацию нуклеотидов.
72. 4. Синдром Леша-Нихана (Найхена).
Снижение регенерации пуриновых нуклеотидов
приводит к заболеванию, которое называется синдромом Леша-Нихана (или Найхена).
При снижении регенерации пуриновых нуклеотидов
основания не могут превратиться в нуклеотиды, что приводит:
1) к дефициту пуриновых нуклеотидов и снижению синтеза РНК и белка, особенно в клетках крови и мозга (т.к. в них нет синтеза de novo),
2) к превращению не потраченных на регенерацию оснований в мочевую кислоту
и к отложению ее солей (уратов) в тканях, особенно в почках.
Из-за повреждения почек отложениями уратов
смерть при синдроме Леша-Нихана наступает в детстве.
При синдроме Леша-Нихана есть такие симптомы, как:
олигофрения, агрессивность и самокалечение
(проявляется в том, что ребенок способен отгрызть себе палец).
Причина снижения регенерации пуриновых нуклеотидов при синдроме Леша-Нихана (и причина самого синдрома) –
сильное снижение активности одного из ферментов регенерации
(гуанин/гипоксантин/фосфорибозил-трансферазы – не зубрить)
из-за мутации гена этого фермента,
то есть синдром Леша-Нихана – заболевание наследственное, генетически обусловленное.
Возможно, что умеренное снижение активности этого фермента способствует появлению подагры и умеренному снижению синтеза РНК и белка в клетках головного мозга.
Регенерация пуриновых нуклеотидов
1) препятствует развитию подагры,
2) ликвидирует одну из причин слабоумия и агрессивности.
При регенерации из основания гуанин образуется ГМФ,
а из гипоксантина – ИМФ (из ИМФ – АМФ и ГМФ).
Примечание: пуриновые основания не синтезируются «в чистом виде», но при распаде нуклеотидов образуются.
72. 5. Синтез д е з о к с и /нуклеотидов. Для ДНК. См. п. 35 и 84.
Есть нуклеотиды, у которых нет атома О во 2-м положении рибозы.
Они называются дез/окси/нуклеотидами
и нужны для синтеза из них ДНК.
Снижение синтеза дезокси/нуклеотидов приводит к снижению синтеза ДНК,
что приводит к:
1) снижению деления клеток (пролиферативных процессов)
и к появлению соответствующей клинической картины (дерматиты, анемия, атрофия ЖКТ и т.д.) и
2) к снижению репарации ДНК (накоплению мутаций).
Нуклеотид может превращаться в дезокси/нуклеотид,
когда у него есть два фосфата,
то есть субстратами процесса являются нуклеотиды АДФ, ГДФ, УДФ,
а продуктами – дАДФ, дГДФ, дУДФ.
При превращении нуклеотидов в дезокси/нуклеотиды происходит
присоединение атомов водорода к нуклеотиду,
то есть происходит восстановление нуклеотида,
поэтому ферменты называются (рибо)нуклеотид/редуктазами.
Источниками 2Н для восстановления нуклеотидов являются:
ГЛУТАТИОН (п.56) или БЕЛОК ТИОРЕДОКСИН.
Они способны отдавать атомы водорода своих SH групп.
В молекуле GSH одна группа SH, поэтому для реакции нужны 2 молекулы GSH.
В молекуле тиоредоксина 2 SH группы, поэтому в реакцию вступает 1 молекула тиоредоксина, обозначается как T(SH)2.
После отдачи водорода глутатион и тиоредоксин превращаются в окисленные формы, которые обозначаются так: GSSG и T(S-S).
Схемы реакций восстановления АДФ:
АДФ + GSH ; АДФ + GSSG или
АДФ + T(SH)2 ; дАДФ + T(S-S).
Далее дАДФ превращается в дАТФ (источником фосфата является АТФ).
Вместо буквы А можете подставить буквы Г и У – получится восстановление ГДФ и УДФ.
УДФ превращается в дУДФ для того, чтобы далее получить ТТФ для ДНК.
Схемы реакций восстановления УДФ:
УДФ + GSH ; УДФ + GSSG или
УДФ + T(SH)2 ; дУДФ + T(S-S).
Далее дУДФ превращается в ТТФ (источником фосфата является АТФ).
Схемы реакций восстановления ГДФ:
ГДФ + GSH ; ГДФ + GSSG или
ГДФ + T(SH)2 ; дГДФ + T(S-S).
Далее дАДФ превращается в дАТФ (источником фосфата является АТФ).
GSSG нужно постоянно превращать в GSH,
то есть восстанавливать окисленную форму глутатина.
Иначе снизится синтез ДНК и обезвреживание АФК (см. п.27).
Восстановление глутатиона катализирует глутатион/редуктаза,
источником водорода при этом является НАДФН.
Схема восстановления глутатиона:
GSSG + НАДФН+ + Н+ ; 2GSH + НАДФ+.
T(SS) тоже нужно постоянно превращать в T(SH)2,
то есть восстанавливать окисленную форму тиоредоксина.
Иначе снизится синтез ДНК.
Восстановление тиоредоксина катализирует тиоредоксин/редуктаза,
источником водорода при этом является НАДФН.
Схема восстановления тиоредоксина:
T(SS) + НАДФН+ + Н+ ; T(SH)2 + НАДФ+.
Превращение НАДФ+ в НАДФН происходит в ПФП (п.35),
для которого нужны глюкоза, инсулин, витамин В1; для синтеза НАДФ+ нужен РР.
Для работы глутатион/редуктазы и тиоредоксин/редуктазы
нужен витамин В2 в составе кофактора ФАД в качестве промежуточного переносчика 2Н от НАДФН.
Синтез пиримидиновых нуклеотидов линейный
Синтез пиримидиновых оснований происходит во всех клетках организма. В реакциях синтеза участвует аспарагиновая кислота, глутамин, СО2, затрачивается 2 молекулы АТФ. В отличие от разветвленного синтеза пуринов этот синтез происходит линейно, т.е. пиримидиновые нуклеотиды образуются последовательно, друг за другом.
Условно можно выделить 3 общих этапа синтеза и реакции синтеза УТФ и ЦТФ:
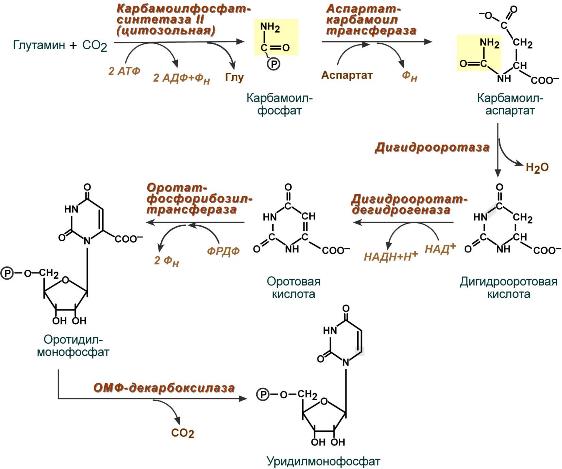
1. Образование карбамоилфосфата
Образование карбамоилфосфата в отличие от синтеза мочевины происходит в цитозоле большинства клеток организма.
2. Образование пиримидинового кольца
3. Синтез оротидинмонофосфата и уридинмонофосфорной кислоты
В реакции с фосфорибозилдифосфатом (ФРДФ) к оротовой кислоте присоединяется рибозо-5-фосфат и образуется оротидилмонофосфат, при декарбоксилировании превращающийся в уридинмонофосфат (УМФ).
Источником фосфорибозилдифосфата является первая из двух реакций синтеза фосфорибозиламина при образовании пуринов.
Синтез уридинмонофосфата
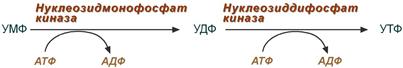
4. Синтез уридинтрифосфата
Синтез УТФ осуществляется из УМФ в 2 стадии посредством переноса макроэргических фосфатных групп от АТФ.
Синтез УТФ
5. Синтез цитидинтрифосфата
Образование цитидинтрифосфата (ЦТФ) происходит из УТФ с затратой энергии АТФ при участии глутамина, являющегося донором NH2-группы.
Какое соединение активирует синтез умф
Фонд пиримидиновых нуклеотидов, подобно пуриновым нуклеотидам, в основном синтезируется из простых предшественников de novo, и только 10 — 20% от общего количества образуется по «запасным» путям из азотистых оснований или нуклеозидов.
А. Образование пиримидиновых нуклеотидов de novo
В отличие от синтеза пуринов, где формирование гетероциклического основания осуществляется на остатке рибозо-5-фосфата, пиримидиновое кольцо синтезируется из простых предшественников: глутамина, СО2 и аспарагиновой кислоты и затем связывается с рибозо-5-фосфатом, полученным от ФРДФ.
Процесс протекает в цитозоле клеток. Синтез ключевого пиримидинового нуклеотида — УМФ идёт с участием 3 ферментов, 2 из которых полифункциональны.
У млекопитающих ключевой, регуляторной реакцией в синтезе пиримидиновых нуклеотидов является синтез карбамоилфосфата из глутамина, СО2 и АТФ, в реакции катализируемой карбамоилфосфатсинтетазой II (КФС II), которая протекает в цитозоле клеток (рис. 10-12). В реакции NH2-rpynna карбамоилфосфата образуется за счёт амидной группы глутамина, что отличает эту реакцию от реакции синтеза карбамоилфосфата в митохондриях в процессе синтеза мочевины из СО2, NH3и АТФ с участием КФС I.
Рис. 10-12. Синтез карбамоилфосфата.

Карбамоилфосфат, использующийся на образование пиримидиновых нуклеотидов, является продуктом полифункционального фермента, который наряду с активностью КФС II содержит каталитические центры аспартаттранскарбамоилазы и дигидрооротазы. Этот фермент назвали «КАД-фермент» — по начальным буквам ферментативных активностей, которыми обладают отдельные каталитические домены этого белка. Объединение первых трёх ферментов метаболического пути в единый полифункциональный комплекс позволяет использовать почти весь синтезированный в первой реакции карбамоилфосфат на взаимодействие с аспартатом и образование карбамоиласпартата, от которого отщепляется вода и образуется циклический продукт — дигидрооротат (рис. 10-13).
Рис. 10-13. Биосинтез УМФ de novo.

Отщепляясь от КАД-фермента, дигидрооротат подвергается дегидрированию NAD-зaвиcимой дигидрооротатдегидрогеназой и превращается в свободное пиримидиновое основание — оротовую кислоту, или оротат.
В цитозоле оротат становится субстратом бифункционального фермента — УМФ-синтазы, которая обнаруживает оротатфосфорибозилтрансферазную и ОМФ-декарбоксилазную активности. Первоначально фосфорибозильный остаток от ФРДФ переносится на оротат и образуется нуклеотид — оротидин-5′-монофосфат (ОМФ), декарбоксилирование которого даёт уридин-5-мо- нофосфат (УМФ).
Таким образом, шесть последовательных реакций синтеза пиримидиновых нуклеотидов осуществляются тремя ферментами, которые кодируются в геноме человека тремя различными структурными генами.
Биосинтез УДФ, УТФ и цитидиловых нуклеотидов
УМФ под действием специфических нуклеозидмонофосфат (НМФ) и нуклеозиддифосфат (НДФ) киназ превращается в УДФ и УТФ в результате переноса у-фосфатного остатка АТФ на соответствующий субстрат.
НМФ-киназа катализирует следующую реакцию:
УМФ + АТФ —> УДФ + АДФ,
УДФ + АТФ —> УТФ + АДФ.
ЦТФ синтетаза катализирует амидирование УТФ (рис. 10-14), осуществляя АТФ-зависимое замещение кетогруппы урацила на амидную группу глутамина с образованием цитидин-5′-трифосфата (ЦТФ).
Рис. 10-14. Синтез ЦТФ из УТФ.

Б. ‹Запасные› пути синтеза пиримидиновых нуклеотидов
Использование пиримидиновых оснований и нуклеозцдов в реакциях реутилизации препятствует катаболизму этих соединений до конечных продуктов с расщеплением пиримидинового кольца. В ресинтезе пиримидинов участвуют некоторые ферменты катаболизма нуклеотидов. Так, уридинфосфорилаза в обратимой реакции может рибозилировать урацил с образованием уридина.
Урацил + Рибозо-1-фосфат —> Уридин + Н3РО4.
Превращение нуклеозидов в нуклеотиды катализирует уридин-цитидинкиназа.
Часть ЦМФ может превращаться в УМФ под действием цитидиндезаминазы и пополнять запасы уридиловых нуклеотидов.
В. Регуляция синтеза пиримидиновых нуклеотидов
Регуляторным ферментом в синтезе пиримидиновых нуклеотидов является полифункциональный КАД-фермент. УМФ и пуриновые нуклеотиды аллостерически ингибируют, а ФРДФ активирует его карбамоилсинтетазную активность, тогда как активность аспартаттранскарбамоилазного домена ингибирует ЦТФ, но активирует АТФ (рис. 10-15).
Рис. 10-15. Регуляция синтеза пиримидиновых нуклеотидов. КАД-фермент катализирует реакции 1,2, 3; дигидрооротатдегидрогеназа — реакцию 4; УМФ синтетаза — реакции 5 и 6; НМФ киназа — реакцию 7; НДФ киназа — реакцию 8; ЦТФ синтетаза — реацию 9.

Этот способ регуляции позволяет предотвратить избыточный синтез не только УМФ, но и всех других пиримидиновых нуклеотидов и обеспечить сбалансированное образование всех четырёх основных пуриновых и пиримидиновых нуклеотидов, необходимых для синтеза РНК.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
© 2018-2021 Все права на дизайн сайта принадлежат С.Є.А.
Биосинтез пиримидиновых нуклеотидов
Механизм синтеза пиримидиновых нуклеотидов почти полностью расшифрован благодаря исследованиям П. Рейхарда. Показано, что в клетках животных и в микроорганизмах конечными продуктами синтеза также не являются свободные пиримидиновые основания и остаток рибозы присоединяется к уже сформировавшемуся пиримидиновому кольцу. Синтез начинается с элементарных уровней (СО2, NH3, аспартат), и специфическую ключевую роль выполняет оротовая кислота.
Последовательность химических реакций синтеза пиримидиновых нуклеотидов, в частности УМФ, можно представить в следующем виде:
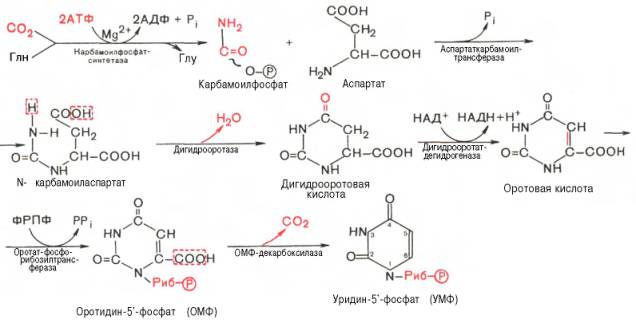
Как видно, I стадия синтеза УМФ включает катализируемое цито-плазматической карбамоилфосфатсинтетазой образование карбамоилфос-фата из глутамина (см. главу 12).
На II стадии карбамоилфосфат реагирует с аспартатом, в результате чего образуется N-карбамоиласпарагиновая кислота. Последняя подвергается циклизации (под действием дигидрооротазы) с отщеплением молекулы воды, при этом образуется дигидрооротовая кислота, которая, подвергаясь дегидрированию, превращается в оротовую кислоту. В этой реакции участвует специфический НАД-содержащий фермент дигидро-оротатдегидрогеназа. Оротовая кислота обратимо реагирует с ФРПФ, являющимся донатором рибозо-фосфата, с образованием оротидин-5′-фос-фата (ОМФ). Декарбоксилирование последнего приводит к образованию первого пиримидинового нуклеотида – уридин-5-фосфата (УМФ).
Превращение УМФ в УДФ и УТФ осуществляется, как и пуриновых нуклеотидов, путем фосфотрансферазных реакций:
Биосинтез цитидиловых нуклеотидов. Предшественником цитидиловых нуклеотидов является УТФ, который превращается в ЦТФ:
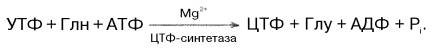
У прокариот в этой реакции используется преимущественно свободный аммиак, в то время как в клетках животных ЦТФ-синтетаза катализирует включение амидной группы глутамина в 4-е положение пиримидинового кольца УТФ. Следует отметить, что образующийся ЦТФ служит отрицательным эффектором регуляторного аллостерического фермента ас-партаткарбамоилтрансферазы, ингибируя по типу обратной связи начальную стадию биосинтеза пиридиновых нуклеотидов. АТФ предотвращает это ингибирование.
Биосинтез тимидиловых нуклеотидов. Тимидиловые нуклеотиды входят в состав ДНК, содержащей дезоксирибозу. Поэтому сначала рассмотрим механизмы синтеза дезоксирибонуклеотидов. При помощи метода меченых атомов было показано, что этот синтез начинается не со свободной дезоксирибозы, а путем прямого восстановления рибонуклеотидов у 2′-го атома углерода. При инкубации меченых предшественников (рибонуклео-тидов) в бесклеточной системе бактерий метку обнаружили в составе дезоксирибонуклеотидов. По данным П. Рейхарда, у Е. coli все 4 рибо-нуклеозиддифосфата восстанавливаются в соответствующие дезоксиана-логи: dАДФ, dГДФ, dЦДФ, dУДФ – при участии сложной ферментной системы, состоящей по меньшей мере из четырех разных ферментов.
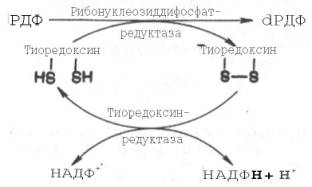
Химический смысл превращения рибонуклеотидов в дезоксирибо-нуклеотиды сводится к элементарному акту – восстановлению рибозы в 2-дезоксирибозу, требующему наличия двух атомов водорода. Непосредственным источником последних оказался восстановленный термостабильный белок тиоредоксин, содержащий две свободные SH-группы на 108 аминокислотных остатков. Тиоредоксин легко окисляется, превращаясь в дисульфидную S-S-форму. Для его восстановления в системе имеется специфический ФАД-содержащий фермент тиоредоксинредуктаза (мол. масса 68000), требующая наличия восстановленного НАДФН. Обозначив условно рибонуклеозиддифосфат РДФ, образование дезоксирибонуклеотидов можно представить следующим образом:
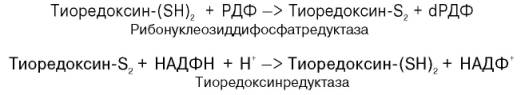
Обе стадии могут быть представлены в виде схемы:
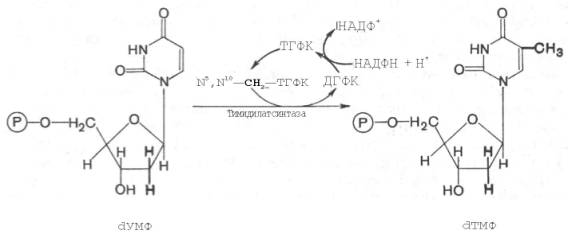
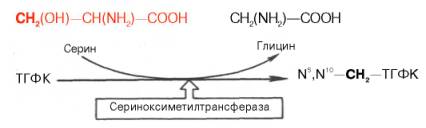
Для синтеза тимидиловых нуклеотидов, помимо дезоксирибозы, требуется также метилированное производное урацила – тимин. Оказалось, что в клетках имеется особый фермент тимидилатсинтаза, катализирующая метилирование не свободного урацила, а dУМФ; реакция протекает по уравнению:
Синтез всех остальных дезоксирибонуклеозид-5′-трифосфатов, непосредственно участвующих в синтезе ДНК, также осуществляется путем фосфорилирования дезоксирибонуклеозид-5′-дифосфатов в присутствии АТФ:
АТФ + dАДФ –> АДФ + dATФ; АТФ + dЦДФ –> АДФ + dЦТФ;
АТФ + dГДФ –> АДФ + dГТФ; АТФ + dТДФ –> АДФ + dТТФ.
Далее на двух схемах суммированы данные о взаимопревращениях пуриновых и пиримидиновых нуклеотидов, а также о связи их с синтезом нуклеиновых кислот. Как видно из схем, в образовании пуриновых и пиримидиновых нуклеотидов специфическое участие принимает ФРПФ, являющийся донором фосфорибозильного остатка в биосинтезе как оро-тидин-5′-фосфата, так и ИМФ; последние считаются ключевыми субстратами в синтезе нуклеиновых кислот в клетках.
Cинтез пиримидиновых нуклеотидов de novo




![]()
![]()
Структура пиримидинового кольца проще, и путь биосинтеза пиримидинов короче, чем у пуринов. Амидный азот глутамина и диоксид углерода обеспечивает атомы 2 и 3 кольца пиримидина после преобразования их в карбамоилфосфат. Другие четыре атома кольца происходят из аспартата. Так же как и в случае с пуриновыми нуклеотидами, углеводная часть поставляется ФРПФ.
Карбамоилфосфат.Образование пиримидиновых нуклеотидов начинается с синтеза карбамоил фосфата, который протекает в цитозоле тканей, способных к образованию пиримидинов (наиболее высокая активность синтеза в селезенке, тимусе, ЖКТ и яичках).
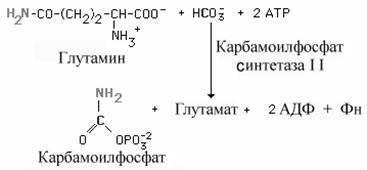 |
Карбамоил фосфат, как упоминалось в главе об обмене аминокислот, используется также в синтезе мочевины.
| Сравнительная характеристика карбомоилфосфат синтетаз I и II | ||
| Карбамоил фосфат синтетаза I | Карбамоил фосфат синтетаза II | |
| Распределение в тканях | Преимущественно печень | Во всех тканях |
| Клеточная локализация | Митохондрия | Цитозоль |
| Метаболический путь | Синтез мочевины | Биосинтез пиримидинов |
| Источник азота | Ионы аммония | Аминогруппа глутамина |
Образование оротовой кислоты. Ключевая реакция.Первая уникальная для биосинтеза пиримидинов реакция – это реакция конденсации карбамоилфосфата и аспартата с образованием карбамоиласпартата, катализируемая аспартаттранскарбамоилазой (реакция
. Реакции синтеза пиримидиновых нуклеотидов
участвующие в синтезе пиримидинов de novo, локализуются в цитозоле. УМФ подвергается двукратному фосфорилированию и образующийся УТФ аминируется с участием глутамина и АТФ и образованием ЦТФ (реакция 9).
Регуляция синтеза пиримидиновых нуклеотидовКлючевым ферментом синтеза пиримидиновых нуклеотидов у человека является цитоплазматическая КФС II. УТФ тормозит активность этого фермента, конкурируя с ATФ. ФРПФ является активатором этого фермента. Имеются и другие участки регуляции (например, OMФ декарбоксилаза ингибируется УМФ и ЦМФ). Однако при нормальных условиях эти участки имеют ограниченное значение. У бактерий ключевым регуляторным ферментом является аспартат транскарбомоилаза. У них имеется только одна карбомоилфосфат синтетаза, так как они не имеют митохондрий. Карбомоилфосфат, таким образом, находится на развилке метаболических путей, которая ведет или к образованию пиримидиновых нуклеотидов или к синтезу аргинина.
Суммарная сравнительная характеристика путей синтеза пуриновых и пиримидиновых нуклеотидов приводится в следующей таблице.



