Сколько камер сердца у кит,уж,крокодил,воробей,лягушка,тритон
![]()
![]()
2) потому что в грибах (даже съедобных) выращенных в лесу могут содержаться токсины молофья и/или содомит серы. у искусственных грибов удаляют абулии, вырабатывающие эти токсины
![]()
![]()
![]()
Комплект молочных зубов состоит из 20 зубов, а комплект постоянных насчитывает — 32.
Все дело в том, что, когда младенец начинает нуждаться в зубах, то есть, когда в его рацион добавляется твердая пища, челюсть малыша еще весьма не больших размеров. Пока ребенок растет, вместе с ним растут и его челюсти. К 6 годам промежутки между зубами становятся довольно таки большими. Это вызвано тем, что челюсти растут, а сами молочные зубы остаются прежних размеров. Уже вскоре после рождения, постоянные зубы начинают формироваться под молочными, которые еще даже не прорезались.
Когда ребенку исполняется 6-7 лет, эти зубы начинают прорезаться. При таком процессе рассасываются корни молочных зубов, молочные зубы и расшатываются. Зуб выпадает, когда укороченный его корень больше не в силах удерживать молочный зуб.
Одна из самых важных функций молочных зубов — это то, что они показывают коренным зубам направление для прорезания. Если молочный зуб выпадает раньше положенного времени (к примеру, есть необходимость в удалении), то постоянный зуб может вырасти криво, ведь он лишился “указателя”.
Видео
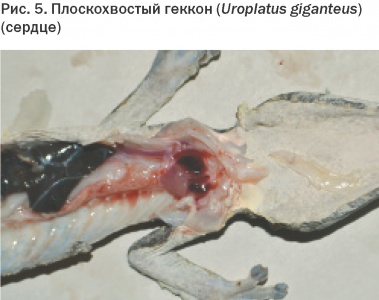
Анатомия и физиология сердечно-сосудистой и дыхательной систем у рептилий. Часть 1
Еще фото
Благодарим редакцию журнал «Мир ветеринарии», Украина, за любезно предоставленную статью Л.А. Стоянова
Анатомия сердечно-сосудистой системы
У рептилий нет одной общей для всех схемы кровообращения. Однако можно выделить два основных типа строения сердца. Первый характерен для чешуйчатых и черепах, а второй – для крокодилов.
Ящерицы, змеи и черепахи
Сердце змей, ящериц и черепах трехкамерное, с двумя предсердиями и одним желудочком (Рис. 1-3). Такое строение предполагает возможность смешивания богатой кислородом крови из легких с кровью, ненасыщенной кислородом, идущей от систем органов. Ряд мышечных гребней и определенная периодичность сокращений служат для функционального разделения желудочка.
Правое предсердие получает ненасыщенную кислородом кровь, которая поступает от всех органов, через венозный синус – расширение на дорсальной стороне предсердия. Стенка венозного синуса мышечная, но не такая толстая, как стенка предсердия. Венозный синус получает кровь от четырех вен:
1. правой передней полой вены;
2. левой передней полой вены;
3. задней полой вены;
4. левой печеночной вены.
Левое предсердие получает насыщенную кислородом кровь из легких через левую и правую легочные вены.
В самом желудочке различают три полости: легочную, венозную и артериальную. Легочная полость – самый вентральный отдел, он продолжается краниально до устья легочной артерии. Артериальная и венозная полости расположены дорсальнее по отношению к легочной и получают кровь от левого и правого предсердия, соответственно. В своей самой краниальной и вентральной части венозная полость дает начало левой и правой дугам аорты (Рис. 4).
Мышечный гребень в некоторой степени отделяет легочную полость от других полостей. Артериальная и венозная полости объединены межжелудочковым каналом.
Одностворчатые атриовентрикулярные клапаны открываются с краниальной стороны межжелудочкового канала. Анатомически они организованы таким образом, что частично закрывают межжелудочковый канал во время систолы предсердия. Во время систолы желудочка их функция заключается в предотвращении регургитации крови из желудочка в предсердия. Серия мышечных сокращений и последующая разница давления в сердце рассматриваемых здесь рептилий разнесены по времени так, чтобы создать функционально двойную систему кровообращения. Систола предсердия нагнетает кровь в желудочек. Расположение атриовентрикулярных клапанов поперек межжелудочкового канала позволяет венозной крови из правого предсердия наполнять венозную и легочную полости. В то же время кровь из легких попадает из левого предсердия в артериальную полость. Желудочковая систола начинается с сокращения венозной полости. Последовательные сокращения венозной и легочной полостей выталкивают кровь из них в малый круг кровообращения с низким давлением.
В продолжение систолы сокращается артериальная полость, что приводит к движению крови через частично сокращенную венозную полость в большой круг кровообращения через левую и правую дуги аорты. Сокращение желудочка приводит мышечный гребень в положение непосредственной близости к вентральной стенке желудочка, создавая таким образом перегородку между артериальной и легочной полостями. Левый и правый атриовентрикулярные клапаны предотвращают возврат крови из желудочка в предсердия.
Все вышеупомянутые явления протекают только при нормальном дыхании. Такая система подачи крови ведет к ее сбросу слева направо на основе разницы давлений. При погружении под воду или в других ситуациях, когда легочное сопротивление и давление повышаются, движение крови происходит справа налево. У красноухой черепахи (Trachemys scripta elegans) при нормальном дыхании кровь движется преимущественно в легочном круге, который получает 60% объема выходящей из сердца крови, а оставшиеся 40% направляются ко всем системам органов. При погружении в воду кровь преимущественно движется по большому кругу, минуя легкие. В таких обстоятельствах давление в легочных сосудах выше, чем на периферии, поэтому кровь входит в сосуды с меньшим давлением – в дуги аорты. У ящериц кровь проходит в основном по левой дуге.
Крокодилы
Строение сердца у крокодилов очень напоминает таковое у птиц и млекопитающих, с той лишь разницей, что у крокодилов есть небольшое отверстие в межжелудочковой перегородке, разделяющей правый и левый желудочки – паниццево отверстие (foramen Pannizi), и что левая дуга аорты выходит из правого желудочка.
Строение сердца крокодилов двойственно по своей природе. Некоторое смешивание насыщенной и ненасыщенной кислородом крови может происходить через паниццево отверстие или в спинной аорте в месте слияния правой и левой дуг. Впрочем, при нормальном дыхании последнего варианта смешивания не происходит, так как давление в системном круге кровообращения превосходит давление в легочном круге. Сброс крови слева направо происходит через паниццево отверстие, и небольшое количество насыщенной кислородом крови попадает в правый желудочек.
Во время погружения под воду или в других условиях, при которых повышается сопротивление легочных сосудов, давление в легочной артерии также существенно возрастает. В результате кровь отводится от легких в системный круг кровообращения. Таким образом, кровь поступает преимущественно в левую дугу аорты, а не в легочную артерию. Существует мнение, что причиной возникновения высокого легочного сопротивления при погружении и, как следствие, сброса крови справа налево, является особый путь оттока крови через правый желудочек. В нем есть отдельная «камера», субпульмонарный конус, который благодаря задержке деполяризации и зубчатым клапанам контролирует поступление крови в легочную сосудистую сеть.
Сам факт сброса крови справа налево при задержке дыхания и повышении сопротивления легочных сосудов может иметь большое клиническое значение. Рептилии под наркозом или без дыхания в отсутствии искусственной вентиляции легких могут демонстрировать непрогнозируемые реакции на ингаляционную анестезию. Кровообращение в обход легких может приводить к недостаточному распределению анестезирующих газов, таких как изофлюран, в системном круге для дальнейших манипуляций под анестезией. Значение длительного сброса справа налево, который может отмечаться при хронических воспалительных процессах в легких, до сих пор мало изучено. При этом можно ожидать серьезных изменений со стороны сердечно-сосудистой системы.
Воротная система почек
Воротная система почек – одна из частей венозной системы рептилий, вызывающая множество вопросов, потенциально имеющих значение для врача. Ее функция заключается в обеспечении достаточного кровоснабжения почечных канальцев при замедлении тока крови через клубочки для сохранения воды.
Афферентные вены воротной системы почек не проникают в клубочки; вместо этого они снабжают кровью проксимальные и дистальные извитые канальцы. Как и у млекопитающих, кровь к клеткам канальцев у рептилий подают приносящие артериолы, которые выходят из клубочков. Однако, в отличие от млекопитающих, в нефронах рептилий нет петель Генле и, следовательно, не происходит реабсорбции воды. В результате, для того чтобы сохранить воду, под воздействием аргининвазо-тоцина замедляется приносящий ток крови через клубочки. При снижении кровоснабжения в клубочках воротная система почек жизненно необходима для подачи крови к канальцам во избежание циркуляторного некроза.
Физиология сердечно-сосудистой системы
Частота сердцебиений у рептилий находится в довольно сложной зависимости от ряда факторов, в том числе температуры тела, размеров тела, уровня обмена веществ, дыхания и внешних раздражителей. Сердечная мышца характеризуется присущей ей максимальной производительностью, измеряемой максимальным напряжением сокращения в пределах зоны оптимальных предпочитаемых температур (ЗОПТ) для данного вида. В общем случае повышение активности приводит к повышению частоты сердцебиений. Частота может увеличиваться втрое по сравнению с частотой сокращений в состоянии покоя. Также, как правило, существует обратная зависимость между размером тела и частотой сердцебиений при заданной температуре.
Интересные вариации частоты сердцебиений при одной и той же температуре окружающей среды проявляются в зависимости от температурного статуса рептилии. В процессе нагревания животное обычно имеет более высокую частоту сердцебиений, чем во время охлаждения. Ускорение сердечного ритма при прогреве помогает достигнуть максимального поглощения тепла. Снижение частоты ударов сердца при понижении температуры окружающей среды помогает рептилии замедлить потерю тепла.
При низких температурах минутный объем сердца, по-видимому, поддерживается за счет увеличения его ударного объема. Ускоренное сердцебиение при повышенных температурах, очевидно, связано со скоростью обмена веществ. Теоретически, высокая частота сердцебиений должна ускорять транспорт кислорода. Изучение кислородного пульса (количество потребляемого кислорода при каждом сердцебиении в мл на массу тела в г) у разных видов говорит об отсутствии последовательной схемы взаимосвязи между объемом сокращения, поглощением кислорода и частотой сердцебиений ввиду повышенной потребности в кислороде при увеличении скорости метаболизма. Различные виды рептилий предположительно обладают множеством механизмов для улучшения подачи кислорода во время ускорения обмена веществ. Отдельно следует упомянуть тот факт, что скорость сердцебиений стремится к увеличению во время активного дыхания и уменьшению при задержке дыхания. Увеличение сердечного ритма совпадает со снижением легочного сопротивления и последующим увеличением легочного кровообращения. Соответственно, увеличение легочного кровообращения в период увеличения дыхательной активности служит для большей эффективности газообмена.
Сердечно-сосудистая система играет ключевую роль в терморегуляции рептилий. Как уже было сказано, частота ударов сердца повышается, когда животное нагревается, и понижается при его охлаждении. Несмотря на то что контролирующий механизм до конца не известен, изменения в кровеносной системе происходят до того, как изменится общая температура тела, что предполагает наличие кожных терморецепторов и барорецепторов.
При нагревании кожных покровов отмечается расширение сосудов в коже. Отток крови в периферические сосуды приводит к падению общего кровяного давления. Понижение сопротивления периферических сосудов способствует развитию сброса крови в сердце справа налево. Кровяное давление поддерживается, таким образом, на уровне, достаточном для снабжения кровью мозга и органов чувств по правой дуге аорты. Кроме того, так как кровь из кожных покровов возвращается в общее кровяное русло, повышается общая температура тела.
Снижение частоты сердцебиений при охлаждении кожи служит для сохранения тепла. При этом наблюдается сужение кровеносных сосудов в коже и относительное расширение сосудов в мышцах. Такое перераспределение крови призвано замедлить теплоотдачу.
Так же как для птиц и млекопитающих, изменения гемодинамики при погружении под воду очень важны и для рептилий. У них имеется ряд преимуществ по сравнению с теплокровными животными, так как рептилии могут использовать альтернативный путь метаболизма при отсутствии кислорода – анаэробный гликолиз. Способность выдерживать анаэробиоз различна у разных видов рептилий. Некоторые ящерицы выдерживают без кислорода не более 25 минут, тогда как некоторые виды черепах способны задерживать дыхание на 33 часа и более. Основные различия заключаются в разной толерантности миокарда к гипоксии.
Как правило, при погружении под воду развивается брадикардия. У крокодилов она обусловлена вагальным торможением сердца под некоторым влиянием торакального или внутрилегочного давления. При нырянии происходит симпатическое сужение кровеносных сосудов в скелетных мышцах, часто до ишемического порога. Такое повышение периферического сопротивления поддерживает кровяное давление для нормальной работы органов.
Сброс крови справа налево возникает при истощении запаса кислорода в паренхиме легких. При дальнейшем погружении сброс справа налево доминирует, практически полностью исключая подачу крови к легким. Общий сердечный выброс может снижаться до уровня в 5% по сравнению с нормальным состоянием. Способность снижать до минимума рабочую нагрузку на сердце, прокачивая лишь небольшую часть крови в системное русло, обеспечивает рептилиям явное преимущество при погружениях по сравнению с птицами и млекопитающими. Брадикардия, связанная с погружением, быстро обратима при первом же вдохе; у некоторых видов даже отмечено ускорение работы сердца еще до выхода на сушу.
Свойства кровеносной системы и их связь с газообменом на клеточном уровне должны приниматься во внимание в любых исследованиях в области кардиологии рептилий. Несмотря на кажущуюся несущественность данного вопроса, клинически подтверждено, что изменения в функционировании сердца или легких могут в значительной степени влиять на способность кровеносной системы к переносу кислорода и углекислого газа.
Молекула гемоглобина считается компонентом, от которого зависят респираторные свойства крови. Хотя структура гемоглобина рептилий пока полностью не описана, она, скорее всего, такая же, как у других позвоночных. Тем не менее, известен ряд существенных отличий в способности гемоглобина удерживать и отдавать кислород. Для этих отличий не было найдено каких-либо закономерностей в зависимости от условий среды, и они не являются общими для всего класса рептилий.
В целом, сродство крови к кислороду зависит от вида рептилии, возраста, размеров и температуры тела. Количество кислорода в организме животного определяется гематокритом и объемом крови. Способность крови переносить кислород зависит от количества эритроцитов на единицу объема (гематокрита). У рептилий он варьирует в пределах: 5-11% у черепах, 6-15% у крокодилов, 8-12% у змей, и от 7% до 8% у ящериц.
Если гемоглобин претерпевает изменения с момента рождения до формирования взрослой особи, то способность крови к насыщению кислородом будет различной в зависимости от этапа онтогенетического развития. При высокой скорости обмена веществ кривые диссоциации кислорода будут смещаться вправо, то есть сродство крови к кислороду будет ниже, что упрощает его доставку к тканям. У рептилий кривые диссоциации кислорода крайне вариабельны. Их сложно обобщить в связи с влиянием непостоянной температуры и скорости метаболизма, а также других перечисленных ранее факторов.
Разные рептилии обладают разными формами гемоглобина, и у некоторых видов гемоглобин эмбриона может иметь сходство к кислороду, отличное от такового у взрослых особей. Гемоглобин может по-разному принимать и отдавать кислород. Эти отличия часто не обнаруживаются клинически, но о них необходимо помнить, чтобы избежать излишней экстраполяции с одного вида на другой.
Сродство к кислороду является мерой того, насколько легко гемоглобин отдает кислород тканям. Гемоглобин с высоким сродством отдает кислород хуже. Низкое сродство означает лучшую отдачу кислорода. У рептилий обычно сродство гемоглобина к кислороду ниже, чем у млекопитающих. Эта адаптация позволяет снабжать кислородом ткани даже при небольшом его содержании в крови.
Во время нагрузок или стресса рептилии могут испытывать метаболический ацидоз вследствие образования молочной кислоты. Изменение рН крови снижает ее сродство к кислороду (эффект Бора), что приводит к тому, что кровь удерживает меньше кислорода и быстрее отдает его тканям.
Изучение кривых диссоциации кислорода у ряда видов рептилий не выявило для них определенных закономерностей. Однако можно предложить несколько общих концепций для отдельных групп рептилий.
Среди ящериц самые активные виды (например, те-йиды, веретеницевые) обладают, как и следовало ожидать, более низким сродством к кислороду. Более высокое сродство к кислороду характерно для медлительных рептилий или для хищников, поджидающих свою добычу (например, хамелеоны, гекконы). Некой серединой для сравнения можно считать игуановых (в том числе, Iguana iguana, Anolis spp., Ctenosaura spp.). Известно, что у игуановых ящериц сродство крови к кислороду напрямую связано с размерами тела. Однако данные, полученные путем измерений при предпочитаемой температуре, слишком ненадежны ввиду поведенческих различий между видами и потому не могут считаться клинически значимыми.
У черепах видимая разница существует между водными и сухопутными видами. Как правило, у водных видов сродство к кислороду ниже, то есть отдача кислорода происходит лучше. У некоторых черепах, живущих в условиях постоянной гипоксии, кровь обладает буферными свойствами, задерживающими эффект Бора, что можно считать адаптацией, связанной с необходимостью максимальной отдачи кислорода во время погружения. Неожиданным исключением является иловая красноватая черепаха (Kinosternum subrubrum), у которой кривая диссоциации кислорода такая же, как и у наземных черепах.
Змеи в этом вопросе принципиально отличаются от черепах. Сравнение водяной яванской бородавчатой змеи (Acrochordus javanicus) и обыкновенного удава (Constrictor constrictor) показало их противоположность по сродству к кислороду. У водяной змеи сродство к кислороду было выше, чем у наземной.
Эта разница может отчасти быть результатом усиленного эффекта Бора, отмечаемого у водных змей. Роль увеличения эффекта Бора, по-видимому, заключается в том, чтобы обеспечить доступность большего количества кислорода в периоды без дыхания при возрастании уровня CO2 в крови. Такая система насыщения крови кислородом позволяет этим видам отдавать кислород, когда это необходимо, во время погружения, и принимать кислород, когда он наиболее доступен, во время дыхательной вентиляции. У змей сродство к кислороду снижается с возрастом, тогда как кислородная емкость (процентный объем кислорода в полностью насыщенной крови) увеличивается по мере роста. Влияние размера тела на сродство к кислороду неодинаково; оно снижается с увеличением размеров (с возрастом) у змей, но повышается у ящериц.
Как и следовало ожидать, кислородная емкость достигает максимума, когда рептилия находится в зоне оптимальных предпочитаемых температур. У змей в связи с нерегулярным типом питания сродство к кислороду падает и его потребление резко возрастает во время переваривания пищи (процесса, требующего усиления обмена веществ). После приема большого количества пищи увеличивается не только потребление кислорода, но и размеры сердца. Андерсон и др. отмечают, что скорость обмена веществ после еды у тигрового питона (Python molurus bivitattus) может увеличиваться до 40%. Высокий уровень метаболизма может сохраняться до 14 дней.
Для поддержания такого уровня обмена сердце питона гипертрофируется на протяжении 48 часов после употребления пищи. Масса сердца может увеличиваться на 40% в ответ на увеличение экспрессии генов сократительных белков мышц. После завершения переваривания пищи размеры сердца возвращаются к норме.
Окончание статьи в следующем номере журнала.
Удавы и питоны слышат, как трепещут сердца жертв.
Недавно ученые выяснили, что змеи не такие уж и глухие, как об этом думали раньше. Оказывается, питоны и удавы могут слышать некоторые звуки. Например, бьется ли еще сердце жертвы, которую они душат, или уже нет. Это помогает определить им, стоит ли дальше тратить силы на то, чтобы сжимать свои смертельные объятия, или можно уже расслабиться.

Уши у змей исчезли, потому что в них сыпалась земля
Представление о том, что змеи практически полностью глухие (если сравнивать их с остальными рептилиями, а так же птицами и млекопитающими), сложилось достаточно давно, еще в позапрошлом веке. Основано оно на том, что у этих существ отсутствует наружное ухо (при этом среднее и внутреннее имеются, однако с внешней средой они напрямую не связаны). Подобные особенности строения герпетологи объясняют адаптацией змей к подземному образу жизни (считается, что их предки большую часть времени проводили под землей). Проще говоря, ушные отверстия исчезли потому, что в них постоянно сыпалась земля.
Как и чем слышат змеи?
Конечно же, полностью глухими змей никто никогда не считал. Эксперименты, проведенные еще в начале ХХ века, показали, что змеи с помощью своих органов слуха могут улавливать вибрации почвы. Поскольку их внутреннее ухо подсоединено к челюстной кости, а она «снимает» вибрации с поверхности земли и передает их внутреннему уху, то что-то змеи все-таки слышать могут.
Танцуют ли змеи под музыку?
Группа зоологов из университета Орхуса (Дания), проведя эксперименты с королевским питоном (Python regius), выяснила, что некоторые «воздушные звуки» змеи все-таки слышат. Они помещали рядом с рептилией динамик, подававший звуки разной частоты, и одновременно с помощью электродов измеряли электрическую активность нейронов мозга в отделах, отвечающих за
(удивительно, но у змей эти отделы находятся примерно там же, где и у нас). Выяснилось, что наибольшую активность нервные клетки питона проявляли в ответ на звуки частотой от 80 до 160 Гц (это аналогично звучанию басовых нот на виолончели). Однако небольшая активность была зафиксирована и при проигрывании звуков другого диапазона.
Звуки змеи слышат без барабанной перепонки
Получается, что змеи все-таки слышат звуки, передаваемые по воздуху. Но каким образом? Ведь барабанной перепонки-то у них нет. Герпетологи, обвешав чувствительными датчиками голову питона, обнаружили, что воздушные колебания вполне способны привести в движение слуховые косточки внутреннего уха. Поэтому можно сказать, что змеи слышат звуки напрямую, без усилителя (роль которого в нашем ухе играет та самая барабанная перепонка).
Слух нужен змеям, чтобы слышать, как бьется сердце жертвы
В то же время другая группа зоологов, работавшая в колледже Дикинсона (США), предположила, что слух необходим удавам и питонам для того, чтобы слышать, бьется ли еще сердце жертвы, которую они душат, или уже нет. И это нужно вовсе не для того, что бы удовлетворить свои «низменные садистские инстинкты» (которых у змей, в отличие от людей, просто нет), а затем, чтобы. не пораниться.
Так вот, американские исследователи предположили, что удав в процессе удушения жертвы, достаточно хорошо слышит, бьется ее сердце или нет. Свою гипотезу они проверили в экспериментах с обыкновенным удавом (Boa constrictor). При этом ученые использовали электронное устройство, имитирующее сердечный ритм. Данный прибор вживлялся мертвой, но еще теплой крысе, после чего эту добычу отдавали удаву.
Также исследователи установили, что удав ощущает биение сердце жертвы не с помощью тактильных рецепторов кожи, а благодаря органам слуха (это подтвердилось показаниями электродов, измеряющих нейронную активность). Причем было установлено, что голова удава в момент «смертельных объятий» часто находилась далеко от грудной клетки жертвы. Возникает предположение, что змеи слышали именно звук, передававшийся по воздуху, а не «ловили» с помощью челюсти вибрацию, идущую по телу жертвы.
Ориентация на сердцебиение жертвы у удавов врожденная
Также зоологам удалось выяснить, что способ ориентироваться на сердцебиение жертвы является для удавов врожденным. Принимавшие участие в эксперименте молодые особи Boa constrictor, появившиеся на свет в неволе, во время удушения также прислушивались к сердцу жертвы, хотя у них и не было еще подобного опыта. Отличие от диких особей было лишь в том, что те душили крыс дольше и сильнее (то есть оно, грубо говоря, количественное, но не качественное).
Добавьте «Правду.Ру» в свои источники в Яндекс.Новости или News.Google, либо Яндекс.Дзен
Быстрые новости в Telegram-канале Правды.Ру. Не забудьте подписаться, чтоб быть в курсе событий.



