Физиология и нарушения водно-солевого обмена (методические материалы к практическим и семинарским занятиям)
Информация
Справочное пособие содержит информацию о физиологии водно-солевого обмена (ВСО). Также представлена информация о методах клинической и лабораторной диагностики нарушений ВСО. Перечислены варианты дисгидрий и методы лечения. Предназначается для врачей всех специальностей, курсантов ФПК и студентов медвузов.
Вода организма

Электролитный состав организма

Факторы, влияющие на перемещение внеклеточной воды в организме
Как уже упоминалось выше, вода является транспортной средой, переносящей питательные вещества и кислород к клеткам и уносящей продукты метаболизма от клеток через интерстициальное пространство в кровоток. Возникает вопрос – каким образом вода «знает» куда и что переносить?
Физиология рассматривает три фактора, определяющих целенаправленное движение воды при транскапиллярном обмене:
2. Часть осмотического давления, создаваемую в биологических жидкостях белками, называют коллоидно-осмотическим (онкотическим) давлением (КОД).
Оно составляет примерно 0,7% осмотического давления (или осмотической концентрации), т. е. около 25 мм рт. ст. (2 мосмоль/кг), но имеет исключительно большое функциональное значение в связи с высокой гидрофильностью белков и неспособностью их свободно проходить через полупроницаемые биологические мембраны.

Механизмы поддержания внутриклеточного объема жидкости и внутриклеточного ионного состава
«Натриевый насос». Мембранная проницаемость Na+ в общем в 10-20 раз меньше, чем К+. Однако наличие градиента концентраций Na+ во вне- и внутриклеточном пространствах и отрицательный внутриклеточный заряд могли бы обеспечить силу, способную двигать Na+ в сторону клетки.
В действительности этого не происходит, поскольку такая сила оказывается сбалансированной другой, действующей в обратном направлении и называемой натриевым насосом. Энергия натриевого насоса, являющегося специфическим свойством клеточной мембраны, обеспечивается гидролизом аденозинтрифосфата (АТФ) и направлена на выталкивание Na+ из клетки [Whittman R., Wheeler К. Р., 1970].
Эта же энергия способствует движению К+ внутрь клетки. Установлено, что противоположно направленные движения К+ и Na+ осуществляются в пропорции 2:3. По мнению М. W. В. Bradbury (1973), с физиологической точки зрения для К+ этот механизм не столь существен, так как последний в норме обладает высокой способностью проникать через клеточную мембрану. Описанный механизм является основным для обеспечения постоянства концентрации клеточных и внеклеточных компонентов. Принципиально важен тот момент, что осмолярность внутриклеточной воды величина достаточно постоянная и не зависящая от осмолярности внеклеточного пространства. Это постоянство обеспечивается энергозависимым механизмом.
Биохимия крови
| Сайт: | Образовательный портал МБФ (ВолгГМУ) |
| Курс: | Нарушения метаболизма. Биохимия специализированных тканей. (Для клин.ордов КЛД) |
| Книга: | Биохимия крови |
Оглавление
1. Общие сведения
2. свойства крови
3. Состав крови. Гематокрит

3.1. Плазма крови
Пла́зма кро́ви (от греч. πλάσμα — нечто сформированное, образованное) — жидкая часть крови, в которой взвешены форменные элементы — вторая часть крови. Процентное содержание плазмы в крови составляет 52—61 %. Макроскопически представляет собой однородную несколько мутную (иногда почти прозрачную) желтоватую жидкость, собирающуюся в верхней части сосуда с кровью после осаждения форменных элементов. Гистологически плазма является межклеточным веществом жидкой ткани крови.
Центрифуги-сепараторы выделяют из крови плазму. Плазма крови состоит из воды, в которой растворены вещества — белки (7—8 % от массы плазмы) и другие органические и минеральные соединения. Основными белками плазмы являются альбумины — 55—65 %, α1-глобулины — 2—4 %, α2-глобулины 6—12 %, β-глобулины8 — 12 %, γ-глобулины — 2-4 % и фибриноген — 0,2—0,4 %. В плазме крови растворены также питательные вещества (в частности глюкоза и липиды), гормоны, витамины, ферменты и промежуточные и конечные продукты обмена веществ, а также неорганические вещества.
В среднем 1 литр плазмы человека содержит 900—950 г воды, 65—85 г белка и 20 г низкомолекулярных соединений. Плотность плазмы составляет от 1,025 до 1,029, pH — 7,36—7,44.
Существует обширная практика собирания донорской плазмы крови. Плазма отделяется от эритроцитов центрифугированием с помощью специального аппарата, после чего эритроциты возвращаются донору. Этот процесс называется плазмаферезом.
Плазма с высокой концентрацией тромбоцитов (богатая тромбоцитами плазма) находит все большее применение в медицине в качестве стимулятора заживления и регенерации тканей организма. В настоящее время на её основе разработана многофункциональная медицинская методика, используемая в стоматологии и косметологии.
3.2. Форменные элементы
У взрослого человека форменные элементы крови составляют около 40—50 %, а плазма — 50—60 %. Форменные элементы крови представлены эритроцитами, тромбоцитами и лейкоцитами:
Кровь относится к быстро обновляющимся тканям. Физиологическая регенерация форменных элементов крови осуществляется за счёт разрушения старых клеток и образования новых органами кроветворения. Главным из них у человека и других млекопитающих является костный мозг. У человека красный, или кроветворный, костный мозг расположен в основном в тазовых костях и в длинных трубчатых костях. Основным фильтром крови является селезёнка (красная пульпа), осуществляющая в том числе и иммунологический её контроль (белая пульпа).
4. Биохимия эритроцита
4.1. Транспорт кислорода кровью
4.2. Созревание эритроцита
4.3. Структурно-метаболические особенности эритроцита
Особенности структурной организации мембраны эритроцитов
Эритроцит окружен плазматической мембраной, структура которой хорошо изучена, идентична таковой в других клетках. Цитоплазматическая мембрана эритроцитов включает бислой фосфолипидов, в то время как белки или «плавают» на поверхности мембран, или пронизывают липиды, обеспечивая прочность и вязкость мембран. Площадь мембраны одного эритроцита составляет около 140 мкм2.
На долю белков приходится примерно 49 %, липидов – 44 %, углеводов –7 %. Углеводы химически связаны либо с белками, либо с липидами и образуют соответственно гликопротеиды и гликолипиды.
Важнейшими компонентами мембраны эритроцитов являются липиды, включающие до 48 % холестерина, 17-28 % – фосфотидилхолина, 13-25 % – сфингомиелина и ряд других фосфолипидов.
Фосфотидилхолин мембраны эритроцитов несет нейтральный заряд, практически не вступает в реакции взаимодействия с положительно заряженными каналами Са2+,, обеспечивая тем самым атромбогенность эритроцитов. Благодаря таким свойствам, как текучесть, пластичность, эритроциты способны проходить через капилляры диаметром
Белки мембраны эритроцита делят на периферические и интегральные. К периферическим белкам относят спектрин, анкирин, белок 4.1., белок р55, адуцин и др. В группу интегральных белков входит фракция 3, а также гликофорины А, В, С, О, Е. Анкирин образует соединение с р-спектрином. В составе эритроцитов обнаружено около 340 мембранных и 250 растворимых белков.
Пластичность эритроцитов связана с фосфорилированием мембранных белков, особенно белков полосы 4.1.
Белок фракции 4.2. – паллидин обеспечивает связывание спектрин-актин-анкиринового комплекса с фракцией 3, относится к группе трансглутаминазных протеинов.
К числу сократительных белков мембраны эритроцитов относятся р-актин, тропомодулин, строматин и тропомиозин.
Гликофорины – интегральные белки мембраны эритроцитов, определяющие отрицательный заряд, способствующий отталкиванию эритроцитов друг от друг и от эндотелия сосуда.
Протеин 3 – основной белок актинов, регулирующий дефосфорилируемость эритроцита.
Как указывалось выше, мембрана эритроцита представляет собой сложный комплекс, включающий определенным образом организованные липиды, белки и углеводы, которые формируют наружный, средний и внутренний слои эритроцитарной мембраны.
Касаясь пространственного расположения различных химических компонентов эритроцитарной мембраны, следует отметить, что наружный слой образован гликопротеидами с разветвленными комплексами олигосахаридов, которые являются концевыми отделами групповых антигенов крови. Липидным компонентом наружного слоя являются фосфатидилхолин, сфингомиелин и неэстерифицированный холестерин. Липиды наружного слоя мембраны эритроцита играют важную роль в обеспечении постоянства структуры мембраны, избирательности ее проницаемости для различных субстратов и ионов. Вместе с фосфолипидами холестерин регулирует активность мембранно-связанных ферментов путем изменения вязкости мембраны, а также участвует в модификации вторичной структуры ферментов. Молярное отношение холестерин / фосфолипиды в мембранах клеток у человека и многих млекопитающих равно 0,9. Изменение этого соотношения в сторону увеличения наблюдается в пожилом возрасте, а также при некоторых заболеваниях, связанных с нарушением холестеринового обмена.
Снижение текучести мембраны эритроцита и изменение ее свойств отмечается также и при увеличении содержания сфингомиелина,
Средний бислой мембраны эритроцита представлен гидрофобными «хвостами» полярных липидов. Липидный бислой обладает выраженной текучестью, которая обеспечивается определенным соотношением между насыщенными и ненасыщенными жирными кислотами гидрофобной части бислоя. Интегральные белки, к которым относятся ферменты, рецепторы, транспортные белки, обладают активностью только в том случае, если находятся в гидрофобной части бислоя, где они приобретают необходимую для активности пространственную конфигурацию. Поэтому любые изменения в составе липидов эритроцитарной мембраны сопровождаются изменением ее текучести и нарушением работы интегральных белков.
Внутренний слой мембраны эритроцита, обращенный к цитоплазме, состоит из белков спектрина и актина. Спектрин является специфическим белком эритроцитов, его гибкие вытянутые молекулы, связываясь с микрофиламентами актина и липидами внутренней поверхности мембраны, формируют своеобразный скелет эритроцита. Небольшой процент липидов во внутреннем слое мембраны эритроцита представлен фосфатидилэтаноламином и фосфатидилсерином. От наличия спектрина зависит подвижность белков, удерживающих двойной бисой липидов.
Одним из важных гликопротеинов является гликофорин, содержащийся как на внешней, так и на внутренней поверхностях мембран эритроцитов. Гликофорин в своем составе содержит большое количество сиаловой кислоты и обладает значительным отрицательным зарядом. В мембране он располагается неравномерно, образует выступающие из мембраны участки, которые являются носителями иммунологических детерминант.
Строение и состояние эритроцитарной мембраны, низкая вязкость нормального гемоглобина обеспечивают значительные пластические свойства эритроцитам, благодаря которым эритроцит легко проходит по капиллярам, имеющим вдвое меньший диаметр, чем сама клетка, и может принимать самые разнообразные формы. Другим периферическим мембранным белком эритроцитов является анкирин, образующий соединение с молекулой Р-спектрина.
Функции эритроцитарной мембраны
Мембрана эритроцитов обеспечивает регуляцию электролитного баланса клетки за счет активного энергозависимого транспорта электролитов или пассивной диффузии соединений по осмотическому градиенту.
В мембране эритроцитов имеются ионно-проницаемые каналы для катионов Na+, K+, для O2, CO2, Cl– HCO3–.
Транспорт электролитов через эритроцитарную мембрану и поддержание его мембранного потенциала обеспечивается энергозависимыми Na+, K+, Ca2+ – АТФ-азными системами.
Мембрана эритроцитов хорошо проницаема для воды при участии так называемых белковых и липидных путей, а также анионов, газообразных соединений и плохо проницаема для одновалентных катионов калия и натрия.
Белковый путь трансмембранного переноса воды обеспечивается при участии пронизывающего мембрану эритроцитов белка «полосы 3», а также гликофорина.
Молекулярная природа липидного пути переноса воды через эритроцитарную мембрану практически не изучена. Прохождение молекул небольших гидрофильных неэлектролитов через эритроцитарную мембрану осуществляется также, как и перенос воды, за счет белкового и липидного путей. Перенос мочевины и глицерина через мембрану эритроцита обеспечивается за счет ферментативных реакций.
Перенос органических анионов через эритроцитарную мембрану обеспечивается, как и транспорт неорганических анионов, при участии белка «полосы 3».
Эритроцитарная мембрана обеспечивает активный транспорт глюкозы, кинетика которого обеспечивается зависимостью Михаэлиса-Ментен. Важная роль в транспорте глюкозы через эритроцитарную мембрану отводится полипептиду полосы 4,5 (белки с ММ 55 кД – возможные продукты распада полипептида полосы 3). Высказывается предположение о наличии специфического липидного окружения у белков – переносчиков сахаров в эритроцитарной мембране.
Неравномерное распределение моновалентных катионов в системе эритроцит – плазма крови поддерживается при участии энергозависимой Na+-помпы, осуществляющей трансмембранный обмен ионов Na+ эритроцитов на ионы К+ плазмы крови в соотношении 3:2. Кроме указанного трансмембранного обмена Na+/K+, Na+ помпа осуществляет еще, по крайней мере, четыре транспортных процесса: Na+→ Na+ обмен; K+→K+обмен; одновалентный вход ионов Na+, сопряженный с выходом К+.
Молекулярной основой Na+ помпы является фермент Na+, K+ –АТФ-аза – интегральный белок, прочно связанный с мембранными липидами, состоящий из 2х полипептидных субъединиц с ММ 80-100кД.
Транспортная система имеет 3 центра, связывающих ионов Na+, локализованных на цитоплазматической стороне мембраны. С наружной стороны мембраны на транспортной системе имеется 2 центра связывания ионов К+. Важная роль в поддержании высокой активности фермента отводится мембранным фосфолипидам.
Функционирование Са2+-помпы обеспечивается нуклеотидами, а также макроэргическими соединениями, преимущественно АТФ, ЦТФ, ГТФ, в меньшей степени ГТФ и ЦТФ.
Как в случае Nа+-помпы, функционирование Са2+помпы в эритроцитах связано с проявлениями активности Са2+, Mg2+ –АТФ-азы. В мембране одного эритроцита обнаруживается около 700 молекул Са2+, Mg2+ –АТФ-азы.
Наряду с барьерной и транспортной функциями, мембрана эритроцитов выполняет рецепторную функцию.
Экспериментально доказано наличие на мембране эритроцитов рецепторов к инсулину, эндотелину, церулоплазмину, а2-макроглобулину, α- и β-адренорецепторов. На поверхности эритроцитов находятся рецепторы к фибриногену, обладающие достаточно высокой специфичностью. Эритроциты также несут на мембране рецепторы к гистамину, ТхА2, простациклину.
В мембране эритроцитов обнаруживаются рецепторы для катехоламинов, снижающих подвижность жирных кислот липидов мембран эритроцитов, а также осмотическую устойчивость эритроцитов.
Установлена перестройка структуры мембраны эритроцитов под влиянием низких концентраций инсулина, гормона роста человека, простагландинов группы Е и Е2.
В мембранах эритроцитов высока и ц – АМФ активность. При увеличении концентраций в эритроцитах ц–АМФ ( до 10–6 М) усиливаются процессы фосфорилирования белков, что приводит в свою очередь к изменению степени фосфорилированности и проницаемости мембран эритроцитов для ионов Са2+.
Эритроцитарная мембрана содержит изоантигены различных систем иммунологических реакций, определяющих групповую принадлежность крови человека по этим системам.
4.4. Антигенная структура эритроцитарной мембраны
Эритроцитарная мембрана содержит различные антигены видовой, групповой и индивидуальной специфичности. Различают два вида изоантигенов эритроцитов, определяющих групповую специфичность крови человек – А и В агглютиногены. Соответственно в плазме или сыворотке крови обнаруживаются две разновидности изоантител – агглютинины α и β. В крови человека не содержатся одноименных агглютиногенов и агглютининов. Их встреча и взаимодействие может возникать при переливании несовместимых групп крови, приводить к развитию агглютинации и гемолиза эритроцитов.
Как известно, I (0) группа крови характеризуется отсутствием в эритроцитах агглютиногенов А и В при наличии в плазме или сыворотке крови агглютининов α и β, встречается у 40-50 % людей стран центральной Европы.
II (А) группа крови характеризуется наличием в мембране эритроцитов агглютиногена А, в то время как в плазме крови содержатся агглютинины β. Указанная группа крови распространена у 30–40 % людей.
III (В) группа крови характеризуется наличием агглютиногена В в мембране эритроцитов, а в плазме или сыворотке крови – наличием агглютининов типа α. Эта группа крови имеет место примерно у 10 % населения.
IV (АВ) группа крови характеризуется наличием в мембране эритроцитов фиксированных А и В агглютиногенов, при этом в плазме или сыворотке крови отсутствуют естественные агглютинины α и β. Данная группа крови встречается у 6 % населения.
Генетический контроль антигенной системы А,В,О мембран эритроцитов представлен генами О, Н, А, В, локализованными в длинном плече 9-й пары хромосом.
Агглютинины α и β относятся к классу Ig M, являются естественными антителами, образуются у ребенка на первом году жизни, достигая максимума к 8 – 10 годам.
Второе место среди антигенных свойств мембран эритроцитов по клинической значимости занимает система Rh – Hr. Впервые Резус-фактор был открыт в 1940 году К. Ландштейнером и А. Винером, содержится в эритроцитах у 85 % людей белой расы. У 15 % людей эти эритроцитарные антигены отсутствуют. В настоящее время установлена липопротеидная природа антигенов данной системы, их насчитывается около 20, они образуют различные комбинации в мембране эритроцитов. Наиболее распространенными резусантигенами являются 6 разновидностей: Rh0 (D), rh’ (C), rh’’ (E), Hr0 (d), hr’ (c), hr’’ (e). Наиболее сильным антигеном этой группы является Rh0 (D).
Антитела системы Rh и Hr – антирезусагглютинины являются приобретенными, иммунными, отсутствуют в крови Rh (-) людей с момента рождения, синтезируются при первом переливании Rh (+) крови Rh (-) реципиенту, а также при первой беременности Rh (-) женщины Rh(+) плодом. При первой беременности эти антитела синтезируются медленно в течение нескольких месяцев в небольшом титре, не вызывая серьезных осложнений у матери и плода. При повторном контакте резус-отрицательного человека с резус-положительными эритроцитами возможен резус-конфликт. Антитела системы Rh – Hr относятся к классу Ig G, поэтому они легко проникают через плацентарный барьер, вызывают реакции агглютинации и гемолиза эритроцитов плода, что сопровождается развитием гемолитической желтухи новорожденных. В случае повторного переливания несовместимой по Rh–антигенам крови донора и реципиента может наблюдаться гемотрансфузионный шок.
Какое давление создают белки плазмы крови
4.1. Транспорт кислорода
В сложных механизмах транспорта газов кровью и газообмена в тканях важная роль отводится эритроцитам, ответственным за доставку О2 к различным органам и удаление образующегося в процессе метаболизма СО2.
Эритроцит – безъядерная клетка, лишенная митохондрий, основным источником энергии для эритроцита служит глюкоза, метаболизируемая в гексозомонофосфатном шунте или цикле Эмбдена-Мейергофа. Транспорт О2 обеспечивается в значительной мере гемоглобином, состоящим из белка глобина и гема. Последний представляет собой комплексное соединение железа и порфирина. Глобин представляет собой тетрамер полипептидной цепи. Hb A (HbA) – основной гемоглобин взрослых содержит 2 – альфа и 2 – бета – цепи, Hb A2 – содержит две альфа и две дельта цепи.
Гем состоит из иона железа, встроенного в порфириновое кольцо. Ион железа гема обратимо связывает одну молекулу О2. С одной молекулой Hb максимально связываются 4 молекулы О2 с образованием оксигемоглобина.
Гем может подвергаться не только оксигенации, но и истинному окислению, когда железо становится из двухвалентного трехвалентным. Окисленный гем носит название гематина, а молекула гемоглобина становится метгемоглобином. В крови человека метгемоглобин находится в незначительных количествах, его уровень резко возрастает при отравлениях. Метгемоглобин не способен отдавать кислород тканям.
В норме метгемоглобин составляет менее 3% общего Hb крови. Основная форма транспорта О2 – в виде оксигемоглобина. Кислород транспортируется артериальной кровью не только в связи с гемоглобином, но и в растворенном виде. Принимая во внимание тот факт, что 1 г Hb может связать 1,34 мл О2, кислородная емкость крови в среднем у взрослого человека составляет около 200 мл/л крови. Одним из показателей кислородного транспорта является насыщение артериальной крови О2(Sa O2), равного отношению О2, связанного с Hb, к кислородной емкости крови:
SaO2=O2, связанного с Hb/O2 емкость крови* 100%.
В соответствии с кривой диссоциации оксигемоглобина насыщение артериальной крови кислородом в среднем составляет 97%, в венозной крови – 75%.
PaO2 в артериальной крови около 100 мм. рт. ст., а в венозной – около 40 мм. рт. ст.
Количество растворенного кислорода в крови пропорционально парциальному давлению О2 и коэффициэнту его растворимости.
Последний для О2 составляет 0,0031/100 мл крови/ 1 мм. рт. ст.. Таким образом, 100 мл крови при PaO2, равном 100 мм. рт. ст., содержит менее 0,31 мл O2.
Диссоциация оксигемоглобина в тканях обусловлена главным образом химическими свойствами гемоглобина, а также рядом других факторов – температурой тела, рН среды, р СО2.
При понижении температуры тела наклон кривой диссоциации оксигемоглобина возрастает, а при ее повышении – снижается, и соответственно снижается сродство Hb к О2.
При снижении рН, т.е. при закислении среды, сродство гемоглобина к О2 уменьшается. Увеличение напряжения в крови СО2 также сопровождается снижением сродства Hb к О2 и уплощением кривой диссоциации оксигемоглобина.
Известно, что степень диссоциации оксигемоглобина определяется содержанием в эритроцитах некоторых фосфорорганических соединений, главным из которых является 2,3 – ДФГ (2,3 дифосфоглицерат), а также содержанием в эритроцитах катионов. В случаях развития алкалозов, поглощение О2 в легких увеличивается, но в то же время затрудняется отдача кислорода тканями. При ацидозах наблюдается обратная картина.
4.2.Утилизация кислорода тканями
Тканевое или клеточное дыхание включает три стадии. На первой стадии пируват, аминокислоты и жирные кислоты окисляются до двухуглеродных фрагментов ацетильных групп, входящих в состав ацетилкофермента А. Последние на втором этапе окисления включаются в цикл лимонной кислоты, где происходит образование высокоэнергетических атомов водорода и высвобождение СО2 – конечного продукта окисления органических субстратов. На третьей стадии клеточного дыхания атомы водорода делятся на протоны (Н+) и «высокоэнергетические» электроны, передающиеся по дыхательной цепи на молекулярный О2 и восстанавливающие его до НО2. Перенос электронов сопряжен с запасом энергии в форме АТФ, т.е. с окислительным фосфорилированием (рис.6).
Касаясь патогенеза метаболических сдвигов, свойственных гипоксическим состояниям, следует отметить, что в организме человека более 90% всего потребляемого кислорода восстанавливается с участием цитохромоксидазы митохондрий, и лишь около 10% кислорода метаболизируется в тканях с участием оксигеназ: диоксигеназы и монооксигеназы.
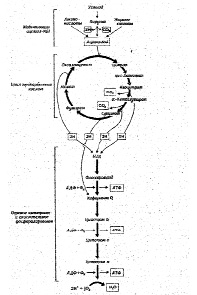
Рис.6. Схема тканевого дыхания. Конечные продукты каждой стадии даны в рамке (Ленинджер А., 1999)
Наиболее многочисленны и сложны монооксигеназные реакции, протекающие в эндоплазматическом ретикулуме клеток при участии цитохрома Р-450 и обеспечивающие гидроксилирование субстрата (стероидных гормонов, лекарственных препаратов и различных др. соединений) и, как правило, его инактивацию.
Диоксигеназы катализируют реакции, в которых в молекулу органического субстрата включаются оба атома молекулы кислорода (например, реакция окисления катехола молекулярным кислородом с раскрытием кольца).
В реакциях, связанных с переносом электронов, т.е. в реакциях окисления-восстановления, где, как указывалось выше, используется более 90% потребляемого кислорода, атомы водорода, отщепленные дегидрогеназами от субстратов в цикле лимонной кислоты, передают свои электроны в цепь переноса электронов и превращаются также в Н +. Как известно, помимо 4 пар атомов водорода, поставляемых каждым оборотом цикла лимонной кислоты, образуются и другие атомы водорода, отщепленные дегидрогеназами от пирувата, жирных кислот и аминокислот в процессе их расщепления до Ацетил-СоА и других продуктов.
Таким образом, все атомы водорода, отщепляемые дегидрогеназами от субстратов, передают свои электроны в дыхательную цепь к конечному акцептору электронов – кислороду.
Скорость утилизации О2 в различных тканях различна. В среднем взрослый человек потребляет 250 мл О2 в 1 мин. Максимальное извлечение О2 из притекающей артериальной крови свойственно миокарду.
Кислород используется в клетках, в основном в метаболизме белков, жиров, углеводов, ксенобиотиков, в окислительно-восстановительных реакциях в различных субклеточных фракциях: в митохондриях, в эндоплазматическом ретикулуме, в реакциях липопероксидации, а также в межклеточном матриксе и в биологических жидкостях.
Коэффициент утилизации О2 в тканях равен отношению потребления О2 к интенсивности его доставки, широко варьирует в различных органах и тканях.
В условиях нормы минимальную потребность в О2 проявляют почки и селезенка, а максимальную потребность – кора головного мозга, миокард и скелетные мышцы, где коэффициент утилизации О2 колеблется от 0,4 до 0,6, а в миокарде до 0,7. При крайне интенсивной физической работе коэффициент утилизации О2 мышцами и миокардом может возрастать до 0,9.
Обмен дыхательных газов в тканях происходит в процессе свободной и облегченной диффузии. При этом О2 переносится по градиенту напряжения газа из эритроцитов и плазмы крови в окружающие ткани.
Одновременно происходит диффузия СО2 из тканей в кровь. На выход О2 из крови в ткани влияет диссоциация оксигемоглобина в эритроцитах, что обеспечивает так называемую облегченную диффузию О2. Интенсивность диффузионного потока О2 и СО2 определяется градиентом их напряжения между кровью и тканями, а также площадью газообмена, плотностью капилляров, распределением кровотока в микроциркуляторном русле. Интенсивность окислительных процессов в тканях определяется величиной критического напряжения О2 в митохондриях, которое в условиях нормы должно превосходить 0,1-1 мм рт. ст.
Соответствие доставки О2 к органам и тканям, возросшим потребностям в оксигенации обеспечивается на клеточном, органном уровнях за счет образования метаболитов изнашивания, а также при участии нервных, гормональных и гуморальных влияний.
Основная масса углекислого газа (СО2) образуется в организме как конечный продукт различных метаболических реакций и транспортируется к легким с кровью. Вдыхаемый воздух содержит лишь незначительное количество СО2.
Транспорт СО2 кровью осуществляется в 3-х состояниях: в виде аниона бикарбоната, в растворенной форме и в виде карбаминовых соединений.
СО2 хорошо растворяется в плазме крови и в артериальной крови, около 5% от общей двуокиси углерода содержится в крови в растворенной форме.
Третьей формой транспорта СО2 кровью являются карбаминовые соединения, образованные взаимодействием СО2 с концевыми группами белков крови преимущественно с гемоглобином:
Карбаминовые соединения составляют около 5% от общего количества СО2, транспортируемого кровью.
В оксигенированной артериальной крови напряжение СО2 составляет 40 мм. рт. ст., а в венозной крови Рv СО2 равно 46 мм. рт. ст.
4.4.Связывание гемоглобина с окисью углерода



